Незрелые половые клетки, достигшие определенной дифференциации, вступают в мейоз.
Мейоз включает два последовательных деления ядра: первое — обычно является редукционным, уменьшающим число хромосом вдвое, второе — эквационным (уравнительным), в основных чертах сходным с митозом. Цикл мейоза состоит из ряда последовательных фаз, в которых хромосомы претерпевают закономерные изменения.
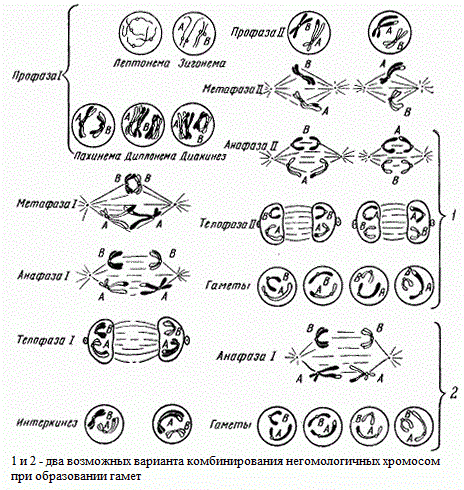
Схема мейоза. А и В — разные пары гомологичных хромосом
Фазы, относящиеся к первому делению, принято обозначать римской цифрой I, а ко второму II:
| Интерфаза | Профаза I | Интеркинез | Профаза II |
| пролептотена | |||
| лептотена | |||
| зиготена | |||
| пахитена | |||
| диплотена | |||
| диакинез | |||
| Метафаза I | Метафаза II | ||
| Анафаза I | Анафаза II | ||
| Телофаза I | Телофаза II |
К редукционному делению относят цикл изменений ядра от профазы I до телофазы I. Затем следует интеркинез — особое состояние клетки между двумя делениями, предшествующее наступлению эквационного деления. Часто телофаза I переходит непосредственно профазу II, минуя интерфазное состояние. Прежде чем перейти к рассмотрению последовательных этапов мейотического деления, отметим основные отличия мейоза от митоза.
Эти отличия особенно значительны в профазе I, когда парные, или гомологичные, хромосомы, из которых одна была привнесена женской, а другая — мужской половой клеткой, соединяются в пары. В митозе подобного процесса нет. В конце профазы и начале метафазы мейоза в экваториальной плоскости располагаются хромосомные объединения, состоящие из двух гомологичных хромосом, называемые бивалентами. В митозе же на экваториальной пластинке мы видим отдельные хромосомы в двойном числе. Следующее очень важное отличие касается анафазы. В редукционном делении в анафазе к полюсам деления отходят гомологичные хромосомы: из каждой пары гомологов одна из хромосом отходит к одному, другая — к другому полюсу; в результате число хромосом в дочерних клетках оказывается уменьшенным вдвое, т. е. становится гаплоидным. В митозе же к полюсам отходят половинки хромосом, число хромосом в дочерних клетках сохраняется таким же, как и в исходной материнской — диплоидным.
Профаза I состоит из ряда последовательных стадий подготовки хромосом к делению. Она начинается с перехода ядра из состояния интерфазы в пролептотенную стадию, за которой следует лептотенная стадия.
Лептотенная стадия характеризуется тем, что сетчатая структура ядра переходит в состояние отдельных тонких нитей, соответствующих хромосомам. Число видимых в световом микроскопе нитей равно диплоидному числу, хромосом. При помощи электронного микроскопа удалось установить, что хромосомные нити лептотенной и даже пролептотенной стадий оказываются как минимум двойными, что означает, что их удвоение происходит еще в интерфазе. Но в силу того, что в этой фазе хромосомы еще не достаточно спирализованы и их половинки тесно примыкают друг к другу, двойственная природа хромосом в световом микроскопе не обнаруживается. Двойственность хромосом и закономерное распределение в них хромомер становятся видимыми по мере развития процесса спирализации в лептотенной стадии. Поскольку хромомеры располагаются всегда в определенных участках гомологичных хромосом, есть основание предполагать, что они являются детерминированными районами начальной спирализации. Лептотенная стадия характеризуется отсутствием притяжения между хромосомными нитями.
Наследующей, зиготенной, стадии две гомологичные хромосомы начинают притягиваться друг к другу. Соединение их в пары чаще начинается с концов или с центромер. Сближение, начавшееся в одной точке, распространяется вдоль хромосомы подобно застежке-молнии. Взаимное притяжение гомологичных хромосом, как мы уже говорили, называют конъюгацией, или синапсисом. Механизм такого притяжения связывают иногда с наличием конъюгационной силы, но какова ее природа, остается невыясненным. Известно лишь, что эта сила действует по всей длине хромосомы и проявляется растянуто во времени соответственно длительности прохождения зиготенной стадии. В настоящее время электронно-микроскопическим методом установлено, что синапсис хромосом сопровождается формированием между ними особой структуры, называемой синаптинемальным комплексом. Расположение хромосом в ядре зиготенной стадии не беспорядочно. В одном случае хромосомы собираются на одной стороне ядра, в другом — распределяются по всему ядру, но их концы почти всегда ориентированы поверхности ядра, к месту, где находится в цитоплазме центросома. Все хромосомы от этого места расходятся радиально и попарно, образуя пучок петель. Такое состояние иногда называют синаптическим букетом.
Важно подчеркнуть, что синапсис проходит лишь между гомологичными хромосомами. Поведение хромосом на этой стадии мейоза принципиально отличается от их поведения в профазе митоза.
С завершением полного синапсиса хромосом ядро вступает в следующую стадию — пахитенную, во время которой дальнейшее усиление приводит к их утолщению. В гомологичных хромосомах этот процесс протекает синхронно. Двойное строение каждой из хромосом теперь становится хорошо различимым: каждая хромосома состоит из двух сестринских хроматид.
Эта стадия удобна для изучения тонкого строения хромосом. Б. Мак-Клинток и другие исследователи обнаружили, что в хромосомах на этой стадии, кроме хромомер, встречаются сильнокрасящиеся узелки. Установлено также, что каждый узелок является постоянным признаком хромосомы, он наследуется и является своеобразным цитологическим маркером (метчиком хромосомы).
На диплотенной стадии происходит перекручивание хромосом и начинают развиваться процессы, обратные, тем, которые имели место в зиготенной стадии. Вместо взаимного притяжения идентичными участками парных хромосом между ними развиваются силы, приводящие к взаимному отталкиванию. Отталкивание центромер друг от друга по принципу одноименно заряженных тел приводит к расхождению хромосом. При расхождении центромер точки соприкосновения несестринских хроматид сползают к дистальным концам хромосом. Вследствие указанных причин образуются Х-образные фигуры, называемые хиазмами. Установлено, что как взаимный контакт (синапсис) между хроматидами, так и образование хиазм имеют прямое отношение к обмену одинаковыми гомологичными участками хроматид, а также к механизму расхождения хромосом в первом и втором делениях мейоза.
Для последней стадии профазы — диакинеза характерно сильное укорочение, а следовательно, и утолщение хроматид. При этом число хиазм уменьшается вследствие перемещения их к концам хромосом (терминализация хиазм). На стадии диакинеза биваленты обособляются, что позволяет даже в случае большого количества хромосом в клетке подсчитать их гаплоидное число. Диакинезом заканчивается профаза I.
Следует подчеркнуть, что в каждой тетраде пара сестринских хроматид сдерживается одной центромерой. Обе центромеры гомологичных хромосом тетрады ориентируются в плоскости экватора, что соответствует метафазе I.
В анафазе I гомологичные хромосомы, каждая из которых содержит пару сестринских хроматид — диады, отталкиваются своими центромерами одна от другой и расходятся к противоположным полюсам. Вследствие этого число их в дочерних ядрах уменьшается ровно вдвое. Важно подчеркнуть, что отцовская и материнская хромосомы каждой пары — униваленты каждого бивалента — могут отходить с равной вероятностью к любому из двух полюсов. При этом хромосомы каждой пары ведут себя по отношению к хромосомам других пар независимо. Поэтому вероятность того, что все отцовские хромосомы отойдут к одному и тому же полюсу, а материнские — к другому, будет равна (1/2)n, где n — соответствует гаплоидному числу хромосом данного организма.
Следует отметить, что хиазмы могут сохраняться вплоть до анафазы I. Хиазмы удерживают вместе родительские хромосомы, образующие биваленты. Это обеспечивает более правильное расхождение обеих центромер каждого бивалента в анафазе I к полюсам веретена. Если бы к моменту анафазы между отцовской и материнской хромосомами не было хиазм, то униваленты гомологичных хромосом могли отходить к одному и тому же полюсу, а это приводило бы к нарушению правильности расхождения отцовских и материнских хромосом.
Расхождение сестринских хроматид каждой исходной хромосомы, как правило, не происходит до второго мейотического деления. Одной из главных причин, препятствующих расхождению сестринских хроматид, является наличие неразделившейся центромеры.
Следующей фазой первого мейотического деления является телофаза I. Продолжительность телофазы сильно варьирует у разных видов. Как правило, телофаза продолжается недолго. С окончанием телофазы I гомологичные хромосомы попадают в разные щетки. Впрочем, следует иметь в виду, что после телофазы 1 не всегда наступает цитокинез иногда он осуществляется лишь после второго мейотического деления, а после первого образуются два гаплоидных ядра в одной клетке.
В интеркинезе в отличие от интерфазы, не происходит удвоения числа хромосом и репликации ДНК. Важно отметить, что сестринские хроматиды перед профазой II оказываются уже удвоенными и состоят из полухроматид. Это указывает на то, что удвоение хромосомных нитей произошло еще в интерфазе.
В метафазе II хромосомы (диады), теперь уже в половинном числе, выстраиваются в экваториальной плоскости, располагаясь центромерами по экватору веретена.
В анафазе II осуществляется разделение центромер, и каждая хроматида унивалента (т. е. диады), становится самостоятельной хромосомой отличие от диады, т. е. хромосомы, состоящей из двух хроматид с одной центромерой, отдельную хромосому анафазы II называют монадой. В анафазе II к полюсам деления расходятся монады.
В телофазе II завершается расхождение хромосом к полюсам и наступает цитокинез.
Итак, в результате первого мейотического деления образуются два ядра с половинным, или гаплоидным, числом хромосом, поэтому первое деление мейоза называют редукционным. Во втором делении каждое дочернее ядро вновь делится, но митотическим путем. Поэтому второе деление называют уравнительным, или эквационным. Следовательно, из каждой клетки, вступившей в мейоз, после двух последовательных делений созревания образуются четыре клетки с половинным числом хромосом.
В качестве примера взята исходная зачатковая клетка с двумя парами негомологичных хромосом (2n = 4). Такое число хромосом имеет, например, растение гаплопаппус (Haplopappus gracilis). В этом случае в анафазе I возможны две комбинации расхождение парных хромосом к полюсам, что ведет к возникновению в телофазе I четырех Типов дочерних ядер, имеющих неодинаковый набор хромосом. Возникновение различных комбинаций (сочетаний) хромосом возможно потому, что расхождение хромосом одной пары (А) происходит независимо от расхождения хромосом другой пары (В). Поэтому в анафазе II возможно образование восьми клеток четырех типов.
Расхождение хромосом одной пары от хромосом другой пары осуществляется независимо от числа хромосом в клетке. Так, если в зачатковой клетке исходное число хромосом 2n = 6, как, например, у растения скерды (Crepis capillaris), то число возможных сочетаний при расхождении хромосом будет равно 23 = 8; при 2n, равном 8, число сочетаний будет 24 = 16. У человека при 2n = 46 число возможных сочетаний равно 223.
Таким образом, в результате мейоза возможны различные сочетания отцовских и материнских хромосом в гаплоидном ядре половой клетки. Это обеспечивает образование у одного организма половых клеток с равным по числу, но различным по составу набором хромосом, что и лежит в основе закономерностей наследования свойств и признаков.
Иногда в силу различных причин появляются организмы с половинным числом хромосом во всех клетках. Такие организмы называются гаплоидами. Размножение гаплоидов возможно либо вегетативно, либо путем партеногенеза (удвоение хромосом происходит в зиготе). Особь, развивающаяся из споры и имеющая половинное число хромосом, называется гаплонтом.
У гаплоида также осуществляется мейоз при развитии половых клеток, но в отличие от диплоида он протекает ненормально и полноценных половых клеток почти не дает. Причиной этого является неравное расхождение хромосом к полюсам в анафазе I. Если гаплоид имеет в соматических клетках n = 12 хромосом, то при их расхождении в анафазе I могут образовываться клетки с числом хромосом в наборе от 0 до 12, где каждая хромосома не имеет гомолога и представляет собой унивалент. Очевидно, что полноценными половыми клетками будут лишь немногие, содержащие 12 хромосом. Неравенство расхождения хромосом в данном случае объясняется Отсутствием синапсиса гомологичных хромосом и независимым их расхождением к полюсам деления. Это и доказывает, что каждая пара хромосом ведет себя независимо от других, негомологичных хромосом в наборе клетки. Подобный пример случайного распределения хромосом в анафазе I можно наблюдать у томатов в процессе мейоза.
Дальше будет видно, что цитологический механизм мейоза представляет собой один из основных материальных процессов, обеспечивающих закономерное наследование свойств и признаков при половом размножении. Описанный мейотический цикл преобразования хромосом включает три основных процесса: 1) синапсис гомологичных хромосом, 2) образование хиазм и 3) расхождение и редукцию числа хромосом. Эти процессы являются общими для абсолютного большинства организмов, но в отдельных случаях они протекают иначе и могут иметь место и в соматических клетках.
Различные отступления от нормальной последовательности процессов мейоза при развитии половых клеток обусловлены главным образом особенностями эволюции способов размножения. Так, например, в процессе образования половых клеток у животных (трутни пчел, наездник) с девственным размножением мейоз замещен митозом. Это вызвано вторичным упрощением мейоза. У некоторых организмов в норме выпадает эквационное деление. Имеется ряд примеров, когда в качестве редукционного деления выступает не первое, а второе мейотическое деление (червецы — Pseudococcus, Phenacoccus).
Для изучения генетики необходимо четко представлять значение митоза и мейоза, их сходство и отличия. В основе мейоза лежит воспроизведение хромосом и их расхождение при делении ядра, поэтому основой мейоза является митоз. Принципиальное отличие митоза от мейоза состоит в следующем:
- в митозе каждый цикл деления ядра связан с одной репродукцией хромосом, в мейозе два деления связаны с одной репродукцией;
- в митозе каждая хромосома репродуцируется, и в анафазе дочерние хромосомы расходятся к полюсам. При этом гомологичные хромосомы ведут себя независимо. В результате деления каждая дочерняя клетка получает полный набор хромосом с одинаковым содержанием генов. В мейозе в профазе каждая пара гомологичных хромосом конъюгирует, и в дочерних ядрах происходит уменьшение числа хромосом ровно вдвое, соответствующее числу бивалентов. При этом каждая пара гомологов расходится независимо от других пар. В тех случаях, когда по различным причинам утрачивается один из гомологов гомологичных хромосом, одиночные хромосомы, не имеющие гомолога (униваленты), расходятся случайно и попадают лишь в одно из дочерних ядер;
- в силу отсутствия конъюгации хромосом в митозе и наличия ее в мейозе в последнем имеет место продолжительная и сложно протекающая профаза.
Мейоз с его стадиями и фазами развития половых клеток является лишь одним из этапов процесса полового размножения; после мейоза наступает этап формирования зрелых половых клеток — гамет. Процесс образования зрелых половых клеток называют гаметогенезом.