Стебель и корень — органы осевые. Одна из основных общих функций их заключается в проведении воды и растворенных в ней органических и минеральных веществ по растению.
У стебля с этой функцией связана еще необходимость поддерживать ассимилирующую массу листьев в наиболее рациональном положении. В связи с этим в стебле и корне особенно хорошо развиты проводящая и механическая системы, состоящие преимущественно из прозенхимных элементов. В листе же, приспособленном для ассимиляции, испарения и газового обмена, преобладающее значение имеют анатомические элементы паренхимного типа. Разумеется, в стебле и корне для отложения запасов и для участия в процессе передвижения различных растворов весьма хорошо развита система паренхимы, но проводящие и механические элементы доминируют и придают специфический характер строению осевых органов. В листьях также есть и проводящая и механическая ткани, но ассимиляционная ткань и вообще паренхима преобладают и придают листовым органам свойственные им специфические черты организации.
Как уже отмечалось выше, лист развивается в меристеме конуса нарастания побега. Зачаток будущего листа закладывается в поверхностных слоях меристемы конуса нарастания в виде бугорка. Образовавшийся вырост или бугорок вначале некоторое время вытягивается, причем в разрастании зачатка листа большое участие принимает группа апикальных клеток, производящая за этот период все новые и новые элементы. Затем у большинства растений верхушечный рост листа прекращается. Лишь у некоторых споровых растений, таких, как папоротники, верхушечный рост листа продолжается сравнительно долго, иногда больше года. Взамен верхушечного роста начинает разрастаться вся листовая пластинка и особенно ее базальная часть, в которой дольше сохраняется меристематическая способность клеток. Вообще же рост листа в отличие от осевых органов продолжается относительно недолго.
Параллельно с ростом идет и дифференциация тканей листа. Лист дифференцируется очень рано: основные черты структуры вполне отчетливо выражены уже в маленьких листовых зачатках. У большинства растений умеренной зоны листья по форме пластинки еще в почках вполне походят на взрослые, отличаясь от них лишь незначительными размерами клеток. В таких случаях рост листа по выходе из почки осуществляется преимущественно за счет растяжения клеток.
У древесных растений умеренной зоны листья быстро вырастают и созревают. У некоторых же тропических растений и у травянистых форм, образующих широкие листья, лист разрастается относительно долго. Линейные листья злаков и некоторых других однодольных растений обладают интеркалярным ростом, напоминающим рост оснований междоузлий соломины злаков. Параллельно разрастанию окончательно дифференцируются клетки мезофилла (мякоти листа) и проводящей системы. Ассимиляционная ткань листа носит название мезофилла. Она состоит из паренхимных клеток и заполняет все пространство в массиве листовой пластинки, заключенное между эпидермисами, исключая элементы проводящей системы и вместилища различных выделений. В листьях большинства растений мезофилл дифференцирован на две хорошо различимые части: верхнюю и нижнюю. Эти комплексы клеток настолько хорошо отличаются друг от друга, что могут быть отнесены к разным тканям.
Верхняя часть мезофилла состоит из удлиненных клеток, плотно сомкнутых между собой, цилиндрической или слегка конической формы, причем длинная ось этих клеток находится в положении, перпендикулярном к поверхности верхнего эпидермиса. Эта ткань называется столбчатой, или палисадной. Нижняя часть мезофилла состоит из клеток, в большинстве случаев рыхло соединяющихся друг с другом и с большими межклетниками между ними. Такую ткань называют губчатой.
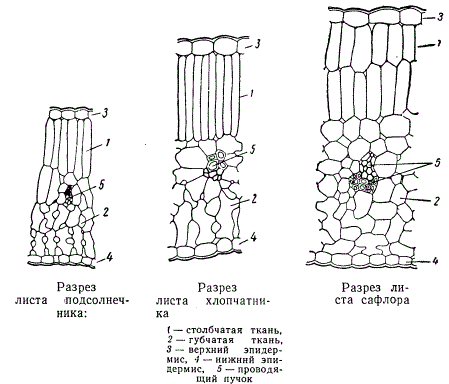
В зависимости от условий произрастания растения, от особенностей его биологии и систематического положения палисадная и губчатая ткани развиваются в различной степени. В листьях растений, приспособленных к произрастанию на ярко освещаемых местах (гелиофиты), палисадная ткань построена из весьма вытянутых клеток. На рисунке изображена часть разреза листа хлопчатника, который произрастает в областях с знойным летом, но требует поливки. У растений, способных выносить засуху, палисадная ткань обычно состоит не менее чем из двух слоев клеток.
Структура листа в значительной степени зависит и от положения листовой пластинки. Листья, горизонтально распростертые, как это бывает у большинства растений, характеризуются обычным расположением палисадной и губчатой тканей, т. е. палисадная ткань обращена к верхней поверхности листа, а губчатая — к нижней. Такое строение пластинки листа называется дорзовентральным. Листья же, растущие под некоторым острым углом к стеблю, имеют палисадную ткань по обеим сторонам листовой пластинки. Таким же типом структуры обладают листья, поставленные ребром, как, например, у некоторых видов эвкалипта на побегах определенного возраста. Листья, характеризующиеся одинаковым строением мезофилла как на морфологически верхней, так и на нижней сторонах пластинки листа, называются изолатеральными.
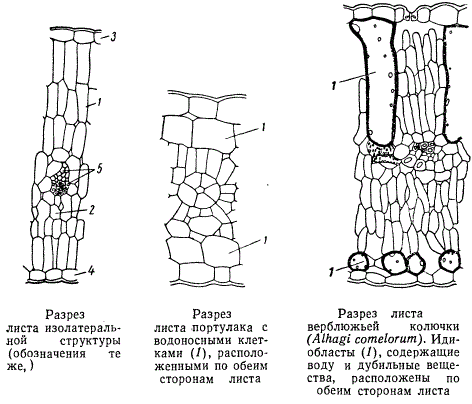
Листья компактной структуры с сильной редукцией системы межклетников и отчетливо выраженной изолатеральностью мезофилла свойственны ксерофитным растениям или вообще растениям, выросшим при недостатке влаги, и потому называются ксероморфными. Листья рыхлой структуры с развитой системой межклетников и подчеркнутой дорзовентральностью в построении мезофилла, свойственные растениям местообитаний с достаточным увлажнением почвы, называются мезоморфными. Изолатеральность свойственна листьям растений не только засушливых мест, но и листьям некоторых растений мезофильного образа жизни. Например, изолатеральны листья ивы и шелковицы, а из травянистых — листья льна.
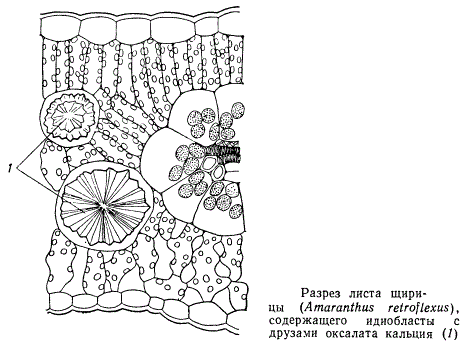
Почти все клетки мезофилла листа содержат хлоропласты и способны к фотосинтезу, но число хлоропластов в различных клетках различно. Больше всего хлоропластов содержится в палисадной ткани, меньше — в губчатой, еще меньше — в так называемых водоносных клетках. При этом пластиды в водоносных клетках часто редуцированы, размеры их весьма малы и хлорофилловая пигментация слабая. Водоносная ткань бывает расположена или с одной стороны листовой пластинки, или с обеих. В листьях некоторых растений засушливых мест, в особенности бобовых, водоносные клетки рассеяны в мезофилле поодиночке. Такие одиночные, достигающие относительно больших размеров клетки, рассеянные среди паренхимы, называются идиобластами. Водоносные идиобласты бобовых содержат в своих вакуолях еще растворы различных дубильных веществ. В большинстве случаев такие идиобласты непосредственно соприкасаются с жилками листа, опираясь на них. Кроме идиобластов, содержащих дубильные вещества, в листьях часто встречаются идиобласты с кристаллическими отложениями оксалата кальция, эфирными маслами и прочими продуктами разносторонней секреторной деятельности растительных организмов.

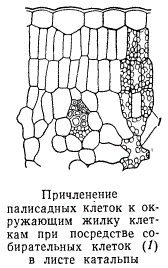
Дифференциация мезофилла листьев на две ткани — палисадную и губчатую — очевидно, способствует наилучшему выполнению одной из основных функций листа. Плотно сомкнутые палисадные клетки своим расположением соответствуют не только направлению падающих на лист солнечных лучей, но и быстрому отведению продуктов ассимиляции. Клетки палисадной ткани или непосредственно опираются на клетки, окружающие жилки листа, или же причленяются к ним с помощью собирательных клеток.
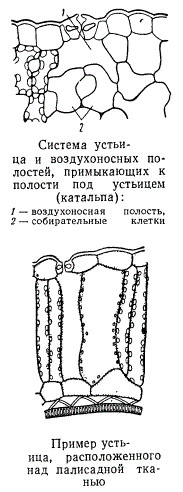
Губчатая ткань листа, имея богатую систему межклетников, наиболее приспособлена к газообмену между листом и окружающей растение средой. На нижней стороне листа у большинства растений образуется больше устьиц, чем на верхней стороне. При этом воздухоносная полость устьица непосредственно сообщается с межклетниками губчатой ткани. У многих растений устьица образуются и на верхней стороне листа и в этих случаях под ними образуются воздухоносные полости.
Пластиды в различных тканях листа различны по своему виду, структуре и по способности накапливать крахмал. Наиболее энергично крахмал накапливается пластидами палисадной и губчатой тканей. При разрастании листа обычно все ткани его растут равномерно. Однако у хвойных эпидермис листьев заканчивает свой рост раньше, чем клетки мезофилла. Вследствие этого клетки мезофилла хвои образуют извилины и складки оболочки. Не следует, однако, полагать, что в листьях хвойных (хвое) совершенно нет межклетников. В хвое вдоль жилки, чередуясь, расположены большие полости, открывающиеся наружу устьицами.
Проводящие ткани листа образуют систему проводящих пучков, или жилок.
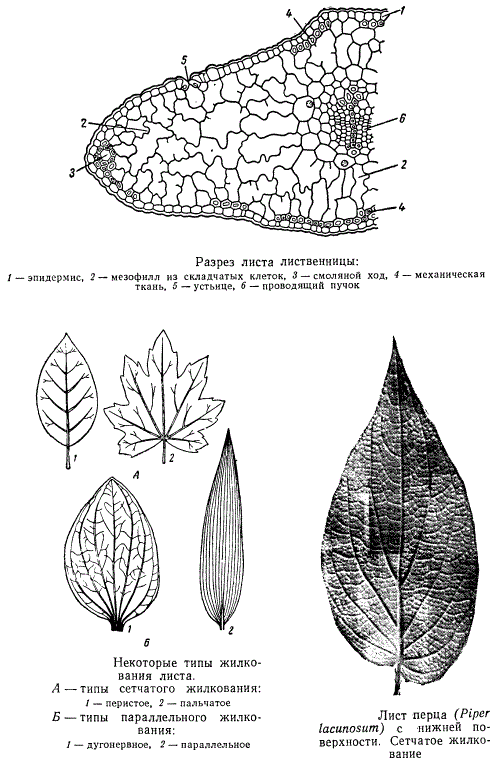
Жилки при рассматривании листа с поверхности обычно видны невооруженным глазом. У покрытосеменных различают два основных типа жилкования: сетчатое и параллельное. В первом случае более мощные жилки постепенно разветвляются на все более тонкие веточки, образующие сеть жилок часто весьма сложного рисунка. При параллельном жилковании вдоль пластинки листа тянутся пучки примерно одинаковой мощности. В верхней части листа и по краю пластинки эти «параллельные» пучки могут сливаться. Кроме того, на всем пути следования они часто соединяются короткими и тонкими анастомозами. Сетчатый тип жилкования свойствен главным образом двудольным, параллельный — однодольным.
Листья многих растений снабжены черешками, но есть немало растений, у которых пластинка листьев доходит до основания листа. Такие листья называются сидячими. Нередко на одном и том же растении нижние листья сидячие, а верхние — черешковые. Первичная проводящая система листовой пластинки и черешка представляет собой одно непрерывное целое с первичной проводящей системой стебля, возникая из общих прокамбиальных тяжей. В листе флоэма обращена к нижней стороне пластинки листа, а ксилема — к верхней стороне. Верхнюю поверхность листа называют еще брюшной, или внутренней стороной, а нижнюю — спинной, или наружной.
В черешке и пластинке листа сосудистые пучки преимущественно коллатерального типа, а у некоторых групп растений — биколлатерального. В черешке пучки или идут отдельными тяжами, или слиты в общую подковообразную или кольцеобразную группу.
У основания черешков, в месте прикрепления их к стеблю, часто расположены прилистники, которые у различных растений бывают неодинаковой величины и формы. Чаще всего прилистники имеют листовидную или чешуевидную форму, а порой достигают таких размеров, что по площади превосходят листовую пластинку (горох); иногда же наоборот, они бывают так малы, что еле заметны, или имеют вид шиловидных придатков у основания черешка (рами, некоторые сорта конопли). В соответствии со степенью развития прилистников проводящая система их либо вполне напоминает проводящую систему листьев, либо сильнейшим образом редуцирована. Проводящая система прилистников сливается с проводящей системой листа часто прежде, чем последняя начнет внедряться в кольцо центрального цилиндра стебля.
Крупные и мелкие жилки в листьях имеют обычно различную структуру: в крупных жилках строение проводящей системы аналогично строению листовых следов в стебле. Наиболее крупные жилки листьев у некоторых растений имеют камбий, который способствует увеличению размеров жилок. Мелкие жилки, в особенности их окончания, часто состоят лишь из одних трахеидоподобных элементов и совершенно не имеют флоэмы.
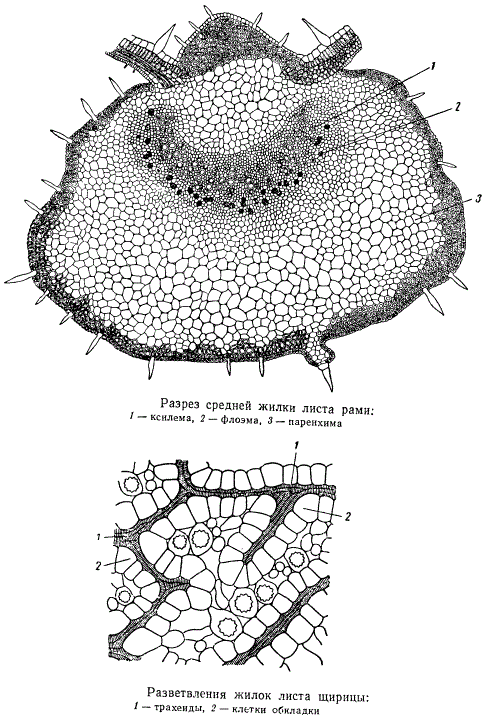
Жилки в листе плотно окружены примыкающими к ним клетками мезофилла. Особенно хорошо такие клетки выражены в листьях ксерофитных растений, т. е. растений, способных выносить значительные периоды засухи. У таких растений клетки, окружающие жилки, нередко бывают настолько крупными и своеобразными, что резко выделяются среди прочей ткани мезофилла. Хлоропластов в них обычно содержится несколько меньше, чем в ассимилирующих клетках. Специфические клетки, окружающие какую-либо группу анатомических элементов, называются обкладочными клетками, а весь комплекс обкладочных клеток — обкладкой. Обкладка принимает большое участие в подъеме и продвижении воды из стебля в лист, а также в процессах передачи продуктов ассимиляции в систему проводящих тканей.
У растений, приспособленных к произрастанию в местах, хорошо снабжаемых водой (мезофиты), клетки обкладок вокруг мелких жилок мало чем отличаются от прочих клеток мезофилла, а у растений, произрастающих в глубокой тени, обкладки иногда нет совсем. В таких случаях к мелким окончаниям жилок примыкают обыкновенные клетки ассимиляционной паренхимы, притом они не облегают жилку сплошь, а оставляют места, где поверхность жилки граничит с межклетниками. Структура обкладки и мощность ее развития прямо зависит от экологических особенностей растения и от его транспирирующей способности.
У многих злаков все жилки листьев снабжены обкладкой из плотно сомкнутых относительно крупных клеток. В определенных местах клетки обкладки одной жилки соединяются с клетками обкладки соседней жилки. Специфические обкладки вокруг жилок присущи не одним только злакам. Они есть у щирицы, портулака и других растений.
Среди культурных злаков хорошо выраженные обкладки характерны для листьев группы просовидных (просо, могар, чумиза и др.), а также для листьев кукурузу, особенно для сортов, произрастающих в засушливых районах. В листьях ржи, ячменя, овса обкладки развиты слабо, несколько сильнее они развиты у пшеницы.
Кроме ассимиляционной и проводящей тканей, в листе есть также система механических тканей. Механические ткани приурочены преимущественно к крупным жилкам. Непосредственно в проводящих пучках развивается твердый дуб. Лубяные волокна нередко образуются и у менее крупных жилок, но, разумеется, не столь мощно. Иногда механические волокна возникают непосредственно в мезофилле листа. По периферии листовой пластинки может формироваться колленхима. Кроме того, у некоторых растений в листьях возникают еще так называемые опорные клетки, одиночно рассеянные в мезофилле листа.
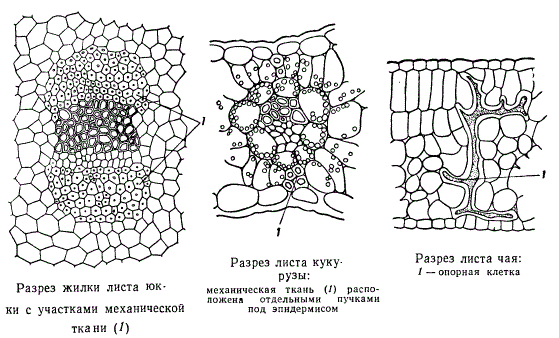
В зависимости от экологического типа растения в листьях по-разному развивается система механических тканей. Листья нежные, тонкие, свойственные растениям тенистых мест или водоемов, имеют незначительное количество механических элементов или даже их нет совсем. Наоборот, растения, произрастающие в местах, подвергающихся интенсивной инсоляции, или в районах с периодически повторяющимися засухами, характеризуются жесткими и колючими листьями, с хорошо развитыми механическими тканями. У растений засушливых мест многие клетки листа, в особенности клетки покровных тканей, склеренхиматизируются, одревесневают, оставаясь морфологически паренхимными. Некоторые растения вообще отличаются усиленным развитием механических тканей в листьях и служат для промышленных целей — изготовления волокна и т. п. Таковы листья некоторых пальм, юкк, драцен, джута и проч. Структура листьев исключительно пластична. На одном и том же растении листья, расположенные на различной высоте одного и того же стебля, отличаются деталями своей структуры, особенно по количественно-анатомическим показателям (закон Заленского). На рисунке показано три листа одного и того же растения (таволги) с частичным анатомическим анализом листьев.
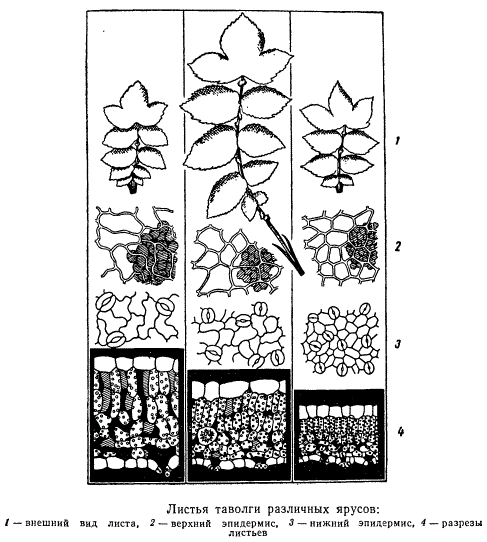
Один лист взят со 2-го яруса, считая от основания стебля, другой — с 10-го яруса и третий — с 18-го яруса. Чем выше место прикрепления листа на стебле, тем мельче становятся его анатомические элементы, больше число их на единицу поверхности листа и теснее смыкание клеток между собой.
Можно вызвать весьма заметные изменения в структуре листьев, особенно изменения количественного характера, и экспериментальным путем. Если высеять растения в различные сроки одного и того же вегетационного периода, то структура листьев таких растений будет различна. Например, в Тбилиси в различное время одного и того же вегетационного периода был высеян подсолнечник одного и того же сорта, выведенного В. С. Пустовойтом: одна группа была посеяна 16-го марта и собрана 6-го июня, другая группа была посеяна 15-го мая и собрана 12-го июля. Кроме того, в каждый срок посадки растения делились на три группы, которые выращивали в различных условиях: одни росли на полном свету и при оптимальной влажности почвы, другие — при уменьшенном содержании воды в почве (40% от полной влагоемкости почвы; оптимальная влажность — 60% от полной влагоемкости), третьи имели достаточно воды, но росли под навесом из белой материи. На рисунке изображены участки поперечных срезов листьев опытных растений. Все срезы получены с листьев одного и того же яруса (7-го). Сравнение рисунков показывает, насколько может меняться структура листьев одного и того же мезофитного растения в различных условиях выращивания.
Вариации в строении листьев можно легко наблюдать и в естественных условиях. Листья, развившиеся в тени, построены значительно более рыхло, чем листья этого же растения, выросшего под прямыми солнечными лучами. Особенно отчетлива разница структуры между световыми и теневыми листьями у древесных растений, например, у сирени, бука, бузины и др.
Листья представляют в большинстве случаев весьма удобный объект для различного рода количественно-анатомических исследований в селекционной работе. Данными количественной анатомии часто пользуются при характеристике сортов какого-либо культурного растения (пшеницы, свеклы и др.) Для этой цели можно использовать как эпидермис, так и мезофилл и жилки листа: измеряют, например, длину устьиц на листьях определенного для всех растений яруса, считая от основания или от верхушки стебля, подсчитывают число устьиц на единицу площади листа, выводят из полученных промеров и подсчетов средние величины и сравнивают результаты, полученные для различных растений. Так же поступают и с клетками мезофилла. Сеть жилок листа на определенной площади срисовывают, а потом курвиметром определяют длину всех жилок, приходящихся на какую-либо единицу площади листа.
Еще шире и разнообразнее применение количественно-анатомических методов при различных экологических исследованиях. Известно, что структура листа отражает результат воздействия различных факторов среды на растения: при недостатке воды или азота листья развиваются мелкоклетными, компактной структуры, а при их избытке листья и межклетники разрастаются. Изменяя в опытах условия произрастания растений и получая ответную реакцию количественного изменения структуры листьев, можно судить как о силе и значении того или другого фактора, так и о степени реагирования растения на воздействия этих факторов.
Количественно-анатомическими методами пользуются также для определения физиологического состояния растения. Измеряя величину щели устьиц, степень открытости ее в тот или другой момент жизни растения, можно судить о том, насколько деятельно общее состояние не только листа, но и всего растения. Изучая состояние устьиц одного и того же растения в различные периоды его вегетации, можно убедиться, что устьица в одно и то же время дня открыты различно: летом устьица открыты не только днем, но иногда и ночью, а осенью, наоборот, закрыты почти все время.
Листья с течением времени стареют, в них развертываются различные дегенерационные процессы: разрушаются пластиды и особенно их пигменты, листья становятся желтыми и буреют. В клетках мезофилла накапливаются различные вещества, которые являются отбросами жизнедеятельности органа.
В некоторых сосудах появляются тиллы, не только закупоривающие сосуд, но и совершенно его разрушающие. Наконец, лист опадает. У некоторых растений он сначала засыхает на растении, а потом отваливается.
Перед опадением листьев в месте прикрепления листа к стеблю, нередко вздутому в виде так называемой листовой подушки, развивается отделяющий слой, состоящий из опробковевших клеток, рассекающий базальную часть черешка поперек. Вещество межклетных пластинок, склеивающее клетки до их опробковения, разрушается, а клетки, подвергнувшиеся опробковению, отмирают и засыхают. Такая ткань, состоящая в значительной мере из отмерших и слабо соединенных между собой клеток, не может противостоять действию тяжести листовой пластинки, и листья, держащиеся на растении почти исключительно при помощи сосудисто-волокнистых пучков, от незначительного сотрясения опадают.
В упавшем на землю листе все клетки постепенно отмирают и разрушаются, причем самыми «живучими» клетками листьев оказываются замыкающие клетки устьиц. Иногда устьица на сильно пожелтевших, уже начавших загнивать участках листьев открыты и содержат пластиды, окончательно не потерявшие своего зеленого цвета.