Основные понятия
Прочная оболочка — характерный признак растительной клетки, отличающий ее от клетки животной.
Оболочка придает клетке определенную форму и прочность и защищает живое содержимое — протопласт и прежде всего плазма-, лемму, плотно прижатую к оболочке изнутри. Клеточные оболочки наземного растения, возвышающегося над поверхностью почвы и не имеющего внутреннего скелета, представляют собой своего рода поддерживающий остов, придающий растению механическую прочность. У каждой клетки есть своя собственная оболочка. Оболочки соседних клеток как бы сцементированы межклетными веществами, образующими так называемую срединную пластинку. Поэтому стенки, разделяющие соседние клетки, в известном смысле двойные.
Оболочка строится живым содержимым клетки и поэтому может расти, только находясь в контакте с ним. Постоянный контакт с протопластом оболочка сохраняет и во взрослой вакуолизированной клетке. Даже в состоянии плазмолиза, когда протопласт отходит от оболочки, связь эта сохраняется благодаря тончайшим нитям, тянущимся от протопласта к оболочке. Очень часто оболочка значительно переживает протопласт, но это обычно не ведет к прекращению существования клетки как структурной единицы, ибо форма клетки, благодаря прочности оболочки, сохраняется. Поэтому растительные клетки и после отмирания живого содержимого могут выполнять важные функции передвижения воды или механической опоры. Многие важные типы клеток в растении — волокна, членики сосудов, пробка и другие специализированные элементы во взрослом состоянии представляют собой одни клеточные оболочки.
Оболочки растительных клеток столь же разнообразны, как и сами клетки. Некоторые оболочки жестки и толсты, другие очень тонки и нежны. По состоянию и строению оболочки часто можно судить о происхождении и функции клеток различных типов. Так, строение ископаемых растений изучают только путем исследования их оболочек. Материал оболочек, состоящий в основном из целлюлозы, широко используется промышленностью. Свойства древесины и бумаги в большой степени зависят от молекулярной структуры оболочки, и знание этой структуры помогает улучшить технологию и качество продукции. Все это послужило причиной того, что развитие, строение, состав и разнообразие оболочек изучены довольно хорошо.
Химический состав
Клеточная оболочка построена из макромолекул высокополимерных веществ, представляющих собой в основном полисахариды. Эти вещества обычно не растворимы в воде и поэтому придают оболочке известную прочность. Важнейшими из них являются целлюлоза, гемицеллюлозы и пектиновые вещества.
Целлюлоза, или клетчатка, является скелетным веществом клеточной оболочки высших растений. Она образует структурный остов, в который погружены вещества двух других типов. Поэтому удаление этих веществ не вызывает значительного изменения формы оболочки и снижения ее механических свойств. Полимерная молекула целлюлозы состоит из большого числа мономеров — остатков глюкозы, связанных между собой в виде цепочки. Число этих остатков в молекуле целлюлозы неодинаково в клеточных оболочках разных растений, что и определяет различные свойства целлюлозы у разных видов. Целлюлоза обладает двойным лучепреломлением, характерным для кристаллов, однако она не типичный кристалл, так как способна к набуханию, а двойное лучепреломление объясняется строгой трехмерной пространственной ориентацией ее молекул, располагающихся параллельно друг другу. Целлюлоза имеет большое промышленное значение, так как все текстильные товары растительного происхождения и бумага состоят из целлюлозы, полученной из клеточных оболочек растений.
Гемицеллюлозы, иногда называемые также полуклетчатками, представляют собой группу полимерных углеводов, в основе которых лежит не глюкоза, как у целлюлозы, а другие мономеры— различные сахара, важнейшими из которых являются ксилоза, манноза и изомер глюкозы, в меньшей степени — арабиноза и галактоза. В состав одной молекулы гемицеллюлозы могут входить остатки сахара не одного типа, а нескольких, например, арабиноза в сочетании с ксилозой, изомер глюкозы в сочетании с маннозой (смешанные гемицеллюлозы). Называются гемицеллюлозы по составу образующих их мономеров с прибавлением окончания «ан». Например, гемицеллюлоза ксилан состоит из остатков ксилозы, маннан — из остатков маннозы, а в состав наиболее важной Гемицеллюлозы древесины хвойных — глюкоманнана — входят остатки глюкозы и маннозы.
Как и для целлюлозы, для гемицеллюлоз характерно нитчатое строение молекул, однако эти молекулы не имеют строгой пространственной ориентации, но и не являются аморфными. Подобно целлюлозе, они выполняют в теле растения механическую функцию, но некоторые из них могут выполнять также функцию запаса, отлагаясь и вновь расходуясь по мере потребности растения (например, в семенах и плодах).
Пектиновые вещества представляют собой продукты полимеризации близкой к углеводам галактуроновой кислоты (полиурониды), а также сахаров арабинозы (арабинаны) и галактозы (галактаны). Как и у гемицеллюлоз, в состав их молекул могут входить несколько мономеров в различных сочетаниях. Пектиновые вещества отличаются от гемицеллюлоз тем, что образуют не нитчатые, а сильно разветвленные молекулы, без всякой пространственной ориентации и поэтому встречаются в клетке, как правило, в аморфном состоянии. Второй особенностью пектиновых веществ является их высокая гидрофильность, набухаемость в воде и способность образовывать гели и слизи. Благодаря присутствию карбоксильных групп (СООН), они способны образовывать не растворимые в воде соли (пектаты), чаще всего с кальцием и магнием. Пектиновые вещества связаны с гемицеллюлозами постепенными, переходами.
В живых клетках оболочка всегда пропитана водой и находится в набухшем состоянии. Особенно много воды (до 80—90%) содержат оболочки молодых клеток. Оболочка более взрослых и особенно стареющих клеток может инкрустироваться различными веществами, например, лигнином, вызывающим ее одревеснение.
Состав веществ, принимающих участие в образовании клеточной оболочки, зависит таким образом от возраста клетки, а также от вида растения, органа и ткани, в которых данная клетка расположена. Так, оболочка клеток волосков на семенах хлопчатника, в массе образующих хлопок, состоит почти из чистой целлюлозы. Древесина сосны и ели, состоящая в основном из мертвых клеток, т. е. фактически из одних клеточных оболочек, включает (от сухого веса) около 40% целлюлозы, 30% гемицеллюлоз и 30% лигнина; в древесине березы и бука при том же количестве целлюлозы содержание гемицеллюлоз повышается до 40%, а содержание лигнина снижается до 20%; пектиновые вещества в древесине составляют доли процента.
Заложение оболочки. Клеточная пластинка
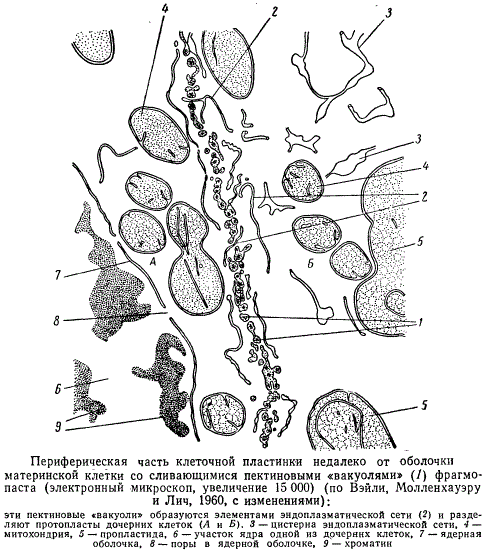
Все эмбриональные клетки, за счет деления которых образуются взрослые специализированные клетки растения, снабжены оболочкой. Поэтому при делении материнской клетки клеточная оболочка должна создаваться заново лишь в плоскости деления. Она и образуется здесь в виде так называемой клеточной пластинки. Как показал световой микроскоп, отложению клеточной пластинки при делении клетки предшествует появление между формирующимися дочерними ядрами волокнистой структуры в цитоплазме, называемой фрагмопластом. Волокна, составляющие фрагмопласт, образуются на основе нитей митотического веретена. Клеточная пластинка закладывается как. свободно плавающая ламелла в центральной части фрагмопласта и развивается в центробежном направлении по средней линии между дочерними ядрами, под прямым углом к волокнам фрагмопласта, постепенно отодвигая их к периферии. Ее формирование связано с возникновением многочисленных мелких пузырьков (пектиновых «вакуолей»), располагающихся в одной плоскости. Эти пузырьки постепенно сливаются в направлении от центра клетки к боковым стенкам, а их содержимое полимеризуетея. В конце концов клеточная пластинка достигает боковых стенок материнской клеточной оболочки и сливается с ними. При этом две образовавшиеся дочерние клетки оказываются разделенными, а фрагмопласт перестает функционировать.
Электронномикроскопические исследования показали, что волокна фрагмопласта представляют собой скопление структур, не отличимых от микротрубочек гиалоплазмы. Роль этих микротрубочек, выявляемых лишь при очень тонких методах фиксации, в делении клетки и образовании клеточной пластинки, как и роль всего фрагмопласта в целом, до сих пор не выяснена. Пузырьки, возникающие на фрагмоплаете и позднее превращающиеся в клеточную пластинку, отчленяются, вероятно, трубчатыми элементами эндоплазматической сети, которая бывает особенно обильной в зоне фрагмопласта. Эти пузырьки- вакуоли вырабатываются и диктиосомами, группирующимися по периферии будущей клеточной пластинки. Они окружены мембраной, которая, вероятно, при слиянии пузырьков принимает участие в образовании плазмалеммы.
Клеточная пластинка образуется на последних этапах деления ядра клетки, и ее формирование обычно заканчивается ко времени расхождения обоих дочерних ядер. Она представляет собой единый слой, общий для двух разделяемых им дочерних клеток. Клеточная пластинка изотропна, так как образована аморфными пектиновыми веществами (главным образом, пектатами кальция и магния), находящимися в состоянии гидрофильного коллоида полужидкой консистенции (геля). Поэтому она очень пластична и способна к значительному растяжению, происходящему при росте дочерних клеток. Во взрослой клетке клеточная пластинка сохраняется, но претерпевает определенные изменения и ее называют поэтому срединной, или межклетной, пластинкой, т. е. пластинкой, общей для двух соседних клеток. Срединная пластинка может быть очень тонкой и в зрелой клетке неразличимой.
Первичная оболочка
Сразу же после образования клеточной пластинки каждый дочерний протопласт начинает откладывать на нее изнутри свою собственную оболочку. При этом отложение оболочки происходит как на поверхности клеточной пластинки, так и на поверхности других клеточных стенок, принадлежащих ранее материнской клетке. Необходимость синтеза нового материала оболочки для уже имеющихся клеточных стенок связана с тем, что после деления клетка вступает в фазу так называемого растяжения. Эта фаза характеризуется очень быстрым увеличением клетки в объеме и ее вакуолизацией. Сохранение непрерывности клеточной оболочки при этом достигается отложением новых порций материала оболочки, синтезируемого протопластом, по всем стенкам клетки.
Рост клетки в фазе растяжения не всегда происходит во все стороны равномерно. Молодые клетки, первоначально обычно изодиаметрические, могут растягиваться преимущественно в одном направлении, при этом возникают цилиндрические и призматические клетки или клетки, вытянутые в виде волокон. Толщина первичной оболочки при этом не уменьшается, так как синтез и отложение оболочки происходят соответственно растяжению клетки.
Вопрос о том, обладает ли первичная оболочка свойствами живого в период растяжения клетки, до сих пор не решен окончательно. Еще недавно предполагали, что в этот период вся первичная оболочка пронизывается цитоплазмой. Однако в электронном микроскопе всегда наблюдается непрерывная плазмалемма, отграничивающая протопласт от оболочки, и не было получено электронограмм, показывающих продолжение цитоплазменных структур за плазмалемму. С другой стороны, оказалось, что эндоплазматическая сеть своими трубчатыми расширениями и диктиосомы тесно связаны с поверхностью клетки в участках растущей оболочки. Следует помнить, однако, что электронный микроскоп улавливает лишь какой-то один момент в жизни клетки и не позволяет изучать поверхность клетки в динамике, что возможно, например, с помощью микрокиносъемки живых клеток. Поэтому вероятно, что в активных клетках плазмалемма принимает неправильные волнистые очертания, а непрерывность ее может нарушаться в связи с прохождением через нее пузырьков, вырабатываемых диктиосомами и эндоплазматической сетью. Материал пузырьков при этом будет откладываться снаружи, а окружающая их мембрана принимать участие в восстановлении непрерывности плазмалеммы. Эти рассуждения лежат в основе более распространенной точки зрения, согласно которой часть цитоплазмы или другие органоиды не входят непосредственно в формирующуюся клеточную оболочку, а выделяют на своей поверхности лишь элементарные единицы (мономеры) и ферменты, необходимые для последующей полимеризации этих структурных веществ и превращения их таким образом в различные полимерные вещества оболочки (например, целлюлозу, пектиновые вещества и др.). Эти элементарные единицы и ферменты и составляют, вероятно, содержимое пузырьков эндоплазматической сети и диктиосом, освобождаемое после их прохождения через оболочку. Об этом свидетельствует и обнаружение в растущей первичной оболочке структурного белка, а ведь все ферменты являются белками.
Первичная оболочка очень тонка и под микроскопом выглядит гомогенной. Она обладает двойным лучепреломлением, так как включает целлюлозу. Однако содержание целлюлозы в первичной оболочке обычно очень невелико (2—12%), и в ней преобладают пектиновые вещества и нерастворимые гемицеллюлозы (ксиланы). В растущей первичной оболочке очень много воды.
Вторичная оболочка

Лубяные волокна льна на поперечном срезе
Отложение первичной оболочки обычно заканчивается, когда клетка перестает расти и достигает своего окончательного размера. Некоторые клетки окружены тонкостенной первичной оболочкой до конца жизни растения. Однако у многих клеток рост оболочки в конце фазы растяжения и в период последующей фазы дифференциации не прекращается, но идет уже в толщину. При этом на внутренней поверхности первичной оболочки откладывается вторичная оболочка. Отложение ее происходит в результате деятельности протопласта, и в мертвых клетках вторичная оболочка не утолщается. Электронномикроскопические исследования показали, что рост вторичной оболочки так же, как и первичной, происходит при включении в нее пузырьков, отчленяемых элементами эндоплазматической сети и диктиосомами, и сопровождается появлением микротрубочек в местах ее синтеза.
Вторичная оболочка может рассматриваться как дополнительная, выполняющая главным образом механическую функцию.
Однако в клетках питательной ткани (эндосперме) некоторых семян вторичная оболочка может выполнять и функцию запаса питательных веществ. В этом случае в ней преобладают запасные гемицеллюлозы, например, маннаны или галактаны. При прорастании семени происходит ферментативный гидролиз этих гемицеллюлоз, вторичная оболочка таких клеток растворяется, и сохраняются только тонкая первичная оболочка и срединная пластинка.
Поэтому толщина ее у разных типов клеток неодинакова и зависит от их специализации. В клетках, выполняющих исключительно опорные, механические функции (каменистые клетки, волокна), вторичная оболочка развита наиболее сильно и может достигать 10 мк толщины. Так как отложение оболочки происходит по направлению к центру клетки (центрипетально), то при этом просвет клетки, занимаемый протопластом и вакуолью, суживается. К моменту достижения клеткой взрослого состояния протопласт может отмирать, а вторичная оболочка достигать такой толщины, что просвет клетки, некогда занятый протопластом, сохраняется лишь в виде небольшой точки в центре клетки.
Довольно часто клетки с сильно развитой вторичной оболочкой теряют живое содержимое и к моменту зрелости становятся мертвыми, состоящими лишь из одних оболочек (некоторые волокна, сосуды). Однако мощную вторичную оболочку могут иметь и клетки с жизнедеятельным протопластом (живые клетки древесины). Чаще же клетки, сохраняющие живой протопласт в период своей физиологической зрелости, имеют, как и эмбриональные клетки, лишь одну первичную оболочку.
При исследовании клеток с достаточно развитой вторичной оболочкой в световом микроскопе часто можно различать три концентрических, различных по мощности слоя: наружный, или переходный, узкий слой, непосредственно прилегающий к первичной оболочке, средний, наиболее мощный и внутренний, узкий слой, граничащий с полостью клетки.
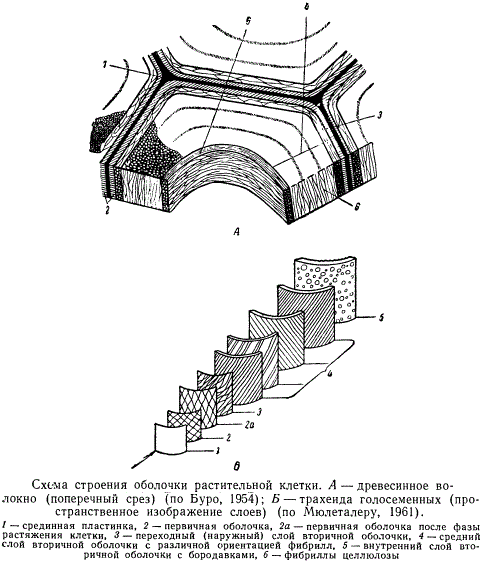
Эти слои отличаются друг от друга по химическому составу (содержанию целлюлозы), физическим свойствам и субмикроскопической структуре. Такое трехслойное строение вторичной оболочки типично. Оно характерно, например, для элементов древесины большинства хвойных. Однако в клетках с менее развитой оболочкой число слоев сокращается до двух за счет выпадения третьего слоя. Иногда же расчленение на слои совсем отсутствует, и вторичная оболочка оказывается совершенно однородной.
Наружный, или переходный, слой вторичной оболочки обычно очень тонкий, отличается присутствием пектиновых веществ. В клетках зрелых тканей он может сливаться почти до полной неразличимости с первичной оболочкой.
Средний, наиболее мощный слой — основной слой, несущий механическую функцию клетки и определяющий свойства вторичной оболочки. Он наиболее богат целлюлозой и не содержит пектиновых веществ. В состав этого слоя может входить и запасная гемицеллюлоза (например, в семенах). Средний слой у многих клеток обладает своеобразной структурой видимой в световой микроскоп в виде слоистости и штриховатости. Слоистость хорошо заметна у более толстых оболочек, например, у лубяных волокон льна и каменистых клеток. Оболочки, имеющие штриховатость, при рассматривании с поверхности кажутся исчерченными очень тонкими косыми штрихами (клетки древесины ели). Слоистость и штриховатость обусловлены субмикроскопической структурой оболочки.
Внутренний слой, называемый часто также третичной оболочкой, очень тонок (500—800 Å) и отличается от среднего слоя иным химическим составом (в частности, повышенным содержанием гемицеллюлоз) и строением. Если содержимое клетки отмирает (например, клетки проводящих и механических тканей), то на поверхности этого слоя образуются многочисленные сферические частицы, получившие название бородавок. Эти бородавки представляют собой, как предполагают, остатки отмершего протопласта. В целом внутренний слой отличается повышенной стойкостью по отношению к веществам, разрушающим клеточную оболочку.
Вторичная оболочка далеко не всегда откладывается равномерно, в виде оплошного слоя. У некоторых специализированных клеток вторичное утолщение ограничено отдельными участками оболочки (местное утолщение). Например, в растении есть клетки, которые, с одной стороны, должны быть прочными, а с другой — проницаемыми для воды. Это клетки, проводящие воду. Во взрослом состоянии они мертвы, лишены тургора и поэтому не имеют давления изнутри. Сохранение ими одновременно известной механической прочности и водопроницаемости достигается за счет того, что вторичная оболочка откладывается не сплошь, а в виде отдельных колец или непрерывной спиральной ленты. Такие клетки, будучи мертвыми, сохраняют способность к вытягиванию, поспевая за ростом соседних, живых клеток. Естественно, что при этом расстояние между кольцами увеличивается.
Структура оболочки
Исследования оболочки с применением электронного микроскопа и различных химических и физических методов позволили установить, что оболочка характеризуется очень тонкой и упорядоченной внутренней структурой. Наличие этой структуры связано со способностью нитчатых молекул целлюлозы располагаться параллельно друг другу и группироваться таким образом, что возникает подобие вытянутой трехмерной решетки, характерной для кристаллов. Такие кристаллические группировки содержат несколько десятков молекул целлюлозы и имеют диаметр 50—70 Å и длину около 600 Å. Их называют обычно элементарными фибриллами, мицеллами, или кристаллитами. Несколько параллельно расположенных элементарных фибрилл собраны, в свою очередь, в нитчатые группы более высокого порядка, называемые микрофибриллами, которые имеют вид прямых палочек, а на продольных срезах — прямоугольников. Плоскость молекул целлюлозы, имеющих форму тяжей, не совпадает с плоскостью слагаемых ими элементарных фибрилл, поэтому эти молекулы показаны на рисунке косыми штрихами.
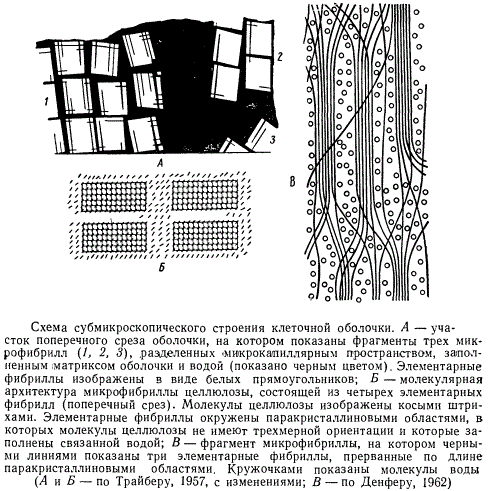
Схема субмикроскопического строения клеточной оболочки
Микрофибриллы представляют собой основные биологические структурные единицы клеточной оболочки. В промежутках между элементарными фибриллами, толщина которых составляет около 10 Å, и по периферии микрофибрилл располагаются молекулы целлюлозы с не столь упорядоченной ориентацией. Эти участки носят название паракристаллических областей микрофибриллы. В состав паракристаллических областей наряду с целлюлозой входят и другие полисахариды (главным образом, гемицеллюлозы).
Размер микрофибрилл колеблется в зависимости от числа составляющих их элементарных фибрилл и равен в среднем 100—250 Å толщины и нескольким тысячам ангстрем длины. Микрофибриллы и элементарные фибриллы видны только в электронном микроскопе.
Отдельные микрофибриллы целлюлозы отделены друг от друга узким миюрокапиллярным пространством порядка 100 Å, заполненным аморфным матриксом (основным веществом) оболочки, состоящим из пектиновых веществ и гемицеллюлоз. В эти же микрокапиллярные пространства откладываются и инкрустирующие вещества, например, лигнин. Микрофибриллы целлюлозы и матрикс оболочки пропитаны водой и находятся в набухшем состоянии. Отдельные элементарные фибриллы связаны в микрофибрилле в единую сеть за счет того, что некоторые из нитчатых молекул целлюлозы переходят от одной элементарной фибриллы к другой, перекрещиваясь и образуя между собой поперечные связи.
Иногда микрофибриллы собраны в группы, образуя пластинки, называемые макрофибриллами, или просто фибриллами. Они представляют собой волокна диаметром около 0,5 мк и длиной несколько микрон, видимые в световой микроскоп и состоящие из большого числа (до 400) параллельно расположенных микрофибрилл, которые лежат в одной плоскости. Капилляры различного размера, разделяющие фибриллы, заполнены матриксом оболочки. Образование фибрилл свойственно главным образом вторичной оболочке.
В первичной оболочке микрофибриллы целлюлозы откладываются в беспорядке в виде рыхлой сети, причем преобладает расположение их перпендикулярно к длинной оси клетки. Положение микрофибрилл может меняться в процессе роста первичной оболочки, так как матрикс в это время еще не стал жесткой структурой. Так как содержание целлюлозы в первичной оболочке невелико, то микрофибрилл в ней сравнительно мало, они не соприкасаются и далеко удалены друг от друга. Основу первичной оболочки в растущей клетке образует аморфный коллоидный материал с высоким содержанием воды и дисперсной фазой из пектиновых веществ и гемицеллюлоз. Такое строение растущей первичной оболочки связано с тем, что в период ее образования (фаза растяжения клетки) она должна обладать пластичностью, а прочности на растяжение в это время не требуется. Поэтому первичная оболочка в фазе растяжения клетки может легко менять форму и быть пластичной, но не эластичной. После прекращения роста клетки, когда начинается отложение вторичной оболочки, содержание воды в первичной оболочке уменьшается, микрофибриллы целлюлозы сближаются друг с другом и теряют способность изменять свое положение. В связи с этим первичная оболочка теряет пластичность и приобретает эластичность.
Во вторичной оболочке, где преобладает уже целлюлоза, а не аморфные вещества, целлюлозные микрофибриллы располагаются более тесно и в правильном порядке, не переплетаясь, и ориентируются обычно параллельно друг другу. Преобладание целлюлозы во вторичной оболочке определяет ее высокие механические качества, особенно прочность на растяжение и эластичность. Фибриллы целлюлозы могут располагаться параллельно или перпендикулярно к продольной оси клетки. В первом случае возникает так называемая поперечная, или кольчатая, текстура вторичной оболочки, во втором случае — продольная, или волокнистая текстура. Кольчатая текстура характерна для клеток, проводящих воду (сосудов). Такая текстура допускает некоторое пластическое (необратимое) растяжение в продольном направлении и одновременно придает оболочке прочность в поперечном направлении, препятствуя сдавливанию клетки с боков. Продольная текстура характерна для клеток, имеющих форму волокон и выполняющих механические функции. Клетки с такой текстурой обладают высокими механическими свойствами в продольном направлении, но могут эластично растягиваться в поперечном направлении. Очень часто целлюлозные фибриллы вторичной оболочки располагаются не строго параллельно продольной оси, а под некоторым углом к ней, в результате чего возникает спиральная текстура оболочки, характерная, например, для лубяных и древесинных волокон и волосков хлопчатника. Причем для наружного и внутреннего слоев вторичной оболочки характерно пологое расположение фибрилл, а для наиболее мощного и наиболее богатого целлюлозой среднего слоя — более крутое их расположение. Клетки с такой текстурой оболочки обладают прочностью как на сжатие, так и на растяжение и изгиб, но в то же время могут эластично (обратимо) растягиваться. Направление фибрилл вторичной оболочки неоднократно меняется под определенным углом к продольной оси клетки, в результате чего и возникает описанная выше слоистость вторичного слоя. Такая различная ориентация фибрилл еще более повышает механические свойства оболочки. Она лежит в основе возникновения штриховатости оболочки, обнаруживаемой при рассматривании ее с поверхности.
Различная ориентация фибрилл связана с характером их образования и отложения в процессе роста оболочки и дифференциации клетки. Еще сравнительно недавно предполагали, что в фазе растяжения клетки рост первичной оболочки происходит путем отложения новых микрофибрилл целлюлозы в промежутки между уже имеющимися микрофибриллами. Такой тип роста, присущий, как считалось, только первичной оболочке, получил название интуссусцепции, или роста внедрением. В основе гипотезы интуюсусцепции лежала концепция проникновения цитоплазмы в растущую оболочку и обнаружение в такой оболочке под электронным микроскопом овальных или округлых участков, свободных от микрофибрилл или с очень рыхлым их расположением. Считалось, что эти участки и представляют собой области разрыхления микрофибрилл. Новый микрофибриллярный материал синтезируется в них, как предполагали, присутствующей здесь цитоплазмой лишь позднее, путем внедрения, или интуссусцепции. При этом межфибриллярная цитоплазма сначала интенсивно растет, а образование микрофибрилл здесь на некоторое время задерживается, что и вызывает возникновение в этих местах участков, свободных от микрофибрилл. Затем в эти участки откладываются новые микрофибриллы. Если таких участков образуется много, то происходит активный поверхностный рост оболочки.
Однако дальнейшие электронномикроскопические исследования показали, что эти участки представляют собой места будущих пор, где синтез оболочки замедлен. Кроме того, как уже говорилось, эти исследования не подтвердили точку зрения о проникновении цитоплазмы в растущую оболочку. Оказалось, что новые микрофибриллы не откладываются в промежутки между старыми, а последовательно накладываются изнутри на уже отложенные микрофибриллы, не переплетаясь с ними и на некотором удалении от них. В результате микрофибриллы, прилегающие к плазмалемме, — самые молодые. Такой способ последовательного отложения слоев микрофибрилл изнутри на уже имеющиеся слон получил название аппозиции, или роста наложением.
В настоящее время считают, что в процессе роста первичной оболочки происходит растяжение не остова из отложенной рыхлой сети микрофибрилл, а пластическое растяжение матрикса. При этом новые микрофибриллы откладываются с внутренней поверхности оболочки преимущественно перпендикулярно к продольной оси клетки способом аппозиции. Затем положение уже отложенных фибрилл может изменяться в связи с отложением изнутри новых микрофибрилл. В результате под действием матрикса происходит пассивная переориентация первоначально параллельно отложенных микрофибрилл, которые в конце концов и образуют в первичной оболочке неправильную рыхлую сеть. Этот процесс повторяется многократно, на протяжении всего периода роста оболочки. Переориентация микрофибрилл возможна в связи с тем, что матрикс оболочки в это время обладает пластичностью. Такой тип роста первичной оболочки носит название многосеточного, так как снаружи оболочка напоминает сеть с крупными ячейками.
Хотя эта теория и объясняет расположение микрофибрилл, однако не дает ответа на вопрос, какие механизмы участвуют в синтезе и поперечной ориентации фибрилл при их выделении цитоплазмой. Природе этих механизмов в настоящее время уделяется большое внимание, особенно в связи с обнаружением микротрубочек в периферической части цитоплазмы клеток, интенсивно синтезирующих оболочку. О механизме синтеза и отложения целлюлозных микрофибрилл существует несколько гипотез, но ни одна из них до сих пор не является общепринятой.
Во вторичной оболочке микрофибриллы также откладываются путем аппозиции, т. е. наложением новых слоев на старые. Но так как вторичная оболочка откладывается тогда, когда клетка уже закончила свой рост, в фазе дифференциации, то первоначальное параллельное расположение микрофибрилл уже не нарушается и последующей переориентации их не происходит.
Несмотря на то, что промежутки между структурным каркасом из целлюлозных микрофибрилл заполнены матриксом из различных веществ наподобие бетона, клеточная оболочка все же является пористой. Об этом свидетельствует прохождение веществ через оболочку. На ранних этапах развития клетки, когда матрикс представляет собой золь — коллоидный раствор с преобладанием воды, различные вещества проходят через оболочку путем диффузии. В дальнейшем, при переходе матрикса в состояние геля в нем, вероятно, остаются незаполненные промежутки различного размера, занятые водой и позволяющие передвигаться даже крупным молекулам веществ.
Срединная пластинка, образующаяся из клеточной пластинки фрагмопласта, первоначально построена почти исключительно из пектиновых веществ. Так как пектиновые вещества аморфны, то срединная пластинка изотропна и не обладает, в отличие от первичной и вторичной оболочек, двойным лучепреломлением. При исследовании -под электронным микроскопом она выглядит совершенно гомогенной.
Мацерация, межклетники
Срединная пластинка как бы цементирует соседние клетки. Поэтому, если ее растворить с помощью некоторых веществ, то оболочки соседних клеток отклеиваются друг от друга, и клетки разъединяются. Процесс разъединения клеток вследствие разрушения срединной пластинки носит название мацерации. Довольно обычна естественная мацерация, при которой пектиновые вещества срединной пластинки переводятся в растворимое состояние с помощью фермента пектиназы и затем вымываются водой. Она наблюдается, например, у перезрелых мясистых плодов груши, дыни и персика. Мучнистый характер плода банана объясняется тем, что ко времени зрелости ткань плода в результате естественной мацерации распадается на отдельные клетки. Естественная мацерация происходит в черешках листьев перед листопадом, в плодоножках и лепестках перед их опадением. В основе процесса мочки льна лежит мацерация, которую производят микроорганизмы. Мацерация наглядно показывает, что каждая клетка обладает своей собственной оболочкой.
При переходе клеток из эмбрионального во взрослое состояние очень часто наблюдается частичная мацерация клеток, при которой срединная пластинка растворяется не по всей поверхности клеток, а лишь в некоторых участках, чаще всего по углам. Вследствие тургорвого давления соседние клетки в этих местах округляются, в результате чего образуются межклетники. Первоначально на поперечном срезе они имеют чаще всего форму трех- или четырехугольников, а на продольном срезе — вид узких щелей. Затем с ростом оболочки соседних клеток межклетники увеличиваются и сливаются, образуя связанную между собой разветвленную сеть. Такая система межклетников улучшает газообмен клеток.
Видоизменения оболочки
Набухшие целлюлозные оболочки очень гибки и эластично растяжимы. Они оказывают очень малое сопротивление диффузии воды и растворенных в ней веществ.
Многие клетки сохраняют целлюлозные оболочки и присущие им свойства без заметных изменений до конца своей жизни. Однако очень часто в процессе развития клетки ее оболочка подвергается вторичным изменениям, приобретая новые химические и физические свойства. Характер этих изменений обычно тесно связан с выполняемыми клеткой функциями. В основе изменений лежит или отложение в оболочку особых веществ или химические изменения уже отложенного вещества. Отложение веществ внутри уже сформированной оболочки (инкрустация) возможно потому, что между микрофибриллами целлюлозы находится система полостей, заполненных водой, которые и служат местом для последующего отложения инкрустирующих веществ. При этом протопласт выделяет, вероятно, не сами вещества, а их предшественников, которые уже на месте, в оболочке, превращаются в инкрустирующие вещества.
Видоизменения оболочки довольно разнообразны. Важнейшими из них являются одревеснение, опробковение, кутинизация, минерализация и ослизнение.
Одревеснение оболочки заключается в инкрустации ее особым веществом — лигнином. Химическая природа лигнина еще окончательно не выяснена. Вероятнее всего он представляет собой не одно вещество, а группу высокополимерных аморфных соединений ароматического ряда. Лигнин откладывается в промежутках вокруг микрофибрилл целлюлозы и превращается таким образом в материал матрикса. При этом он может химически связываться с другими веществами матрикса или с микрофибриллами целлюлозы. Так как целлюлоза обладает большой прочностью на растяжение, а лигнин — на сжатие, то лигнифицированную оболочку можно уподобить по прочности железобетону, в котором роль стальной арматуры выполняют целлюлозные микрофибриллы, а роль бетона — лигнин. Такая оболочка теряет пластичность, но приобретает некоторую хрупкость. В целом она представляет единую жесткую структуру, мало способную к набуханию, так как межфибриллярные промежутки у нее в значительной степени заполняются лигнином. Механическая целостность клеток высшего растения связана с лигнином. Большая прочность на сжатие, вызываемая его отложением в оболочках, позволяет наземному растению развивать массивные структуры.
Хотя лигнифицированные оболочки наиболее характерны для клеток древесины и поэтому называются также одревесневшими, одревесневать могут самые различные ткани, вплоть до лепестков.
Физиологическое значение одревеснения не ограничивается одним увеличением прочности на сжатие. Инкрустация оболочки лигнином, обладающим антисептическими свойствами, защищает, вероятно, клетку от нападения некоторых микроорганизмов. Несмотря на более низкую набухаемость одревесневших оболочек, они более прочно удерживают воду, чем пектоцеллюлозные оболочки. Возможно, этим объясняется значительное одревеснение тканей, проводящих воду, а также тканей растений, произрастающих в условиях недостатка влаги (ксерофитов). Одревеснение обычно происходит в клетках, которые к моменту специализации (во взрослом состоянии) отмирают. Оно предотвращает сдавливание этих клеток, потерявших тургор. Однако это вовсе не значит, что одревеснение препятствует нормальной жизнедеятельности клеток. Например, живые клетки древесины с сильно одревесневшими оболочками обнаруживают высокую физиологическую активность в течение нескольких лет. Обычно одревесневшие оболочки уже не изменяются до конца жизни. Лишь очень редко процесс одревеснения становится обратимым. Например, не вполне зрелые плоды айвы довольно жестки, но при длительном хранении их жесткость резко снижается. Первоначальная жесткость плодов связана с присутствием в них большого количества так называемых каменистых клеток, имеющих толстые и сильно одревесневшие оболочки. При хранении плодов айвы происходит естественное раздревеснение оболочек этих клеток, обусловленное, вероятно, деятельностью ферментов.
Процесс одревеснения клеточной оболочки существует только в живой клетке Отложение лигнина начинается обычно одновременно с началом отложения клеткой вторичной оболочки и происходит прежде всего в первичной оболочке. Затем лигнификация распространяется кнаружи — в срединную пластинку и ковнутри — в развивающуюся вторичную оболочку. В конце концов концентрация лигнина оказывается наибольшей в срединной пластинке, постепенно уменьшаясь по направлению к полости клетки. Например, в древесине ели 60—90% лигнина откладывается в первичной оболочке и срединной пластинке и только 10—40% — во вторичной оболочке, которая по массе преобладает.
Опробковение заключается в отложении в оболочку очень стойкого химического вещества суберина. Суберин — это эфир высокополимерных насыщенных и ненасыщенных жирных кислот и оксикиелот сложного состава и представляет собой аморфное очень гидрофобное соединение. Он откладывается преимущественно во вторичной оболочке в виде одной или нескольких пластинок — ламелл, видимых в световой микроскоп и не связанных с фибриллами целлюлозы. На границе с полостью клетки откладывается чисто целлюлозный слой вторичной оболочки.
Субериновая прослойка может быть непрерывной, одевая всю клетку целиком, или же откладывается только в части оболочки. Опробковевшая оболочка непроницаема для воды и газов, поэтому она особенно сильно развита у клеток пробки, защищающих тело растения снаружи. Клетки пробки к моменту завершения дифференциации отмирают, так как их протопласт со всех сторон окружается воздухонепроницаемой оболочкой. При частичном опробковении оболочки клетка долгое время может оставаться живой.
Кутинизация оболочки состоит в отложении в ней кутина — вещества, очень близкого к суберину, но отличающегося от него низким содержанием ненасыщенных жирных кислот и более высокой степенью полимеризации. Кутин обычно откладывается в оболочку вместе с воском — близким к нему соединением. В отличие от кутина и суберина воск легко извлекается растворителями жиров и легко плавится.
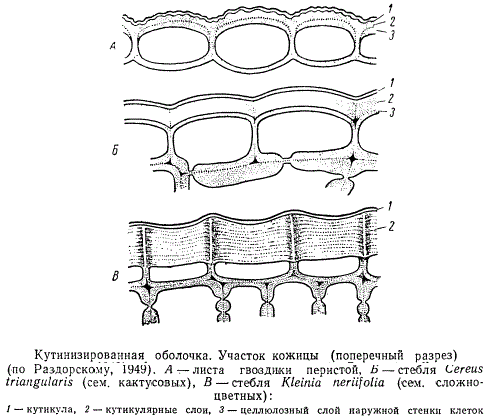
Кутинизированная оболочка клетки
Кутинизации обычно подвергается наружная стенка клеточной оболочки кожицы (покровной ткани), граничащей с атмосферой, иногда кутинизируются и радиальные стенки оболочки этих клеток. В типичном случае кутинизированная оболочка имеет следующее строение. Снаружи откладывается чистый кутан, образуя непрерывный слой кутикулы различной толщины. Далее вглубь следуют так называемые кутикулярные слои оболочки, состоящие из гидрофильных ламелл целлюлозы и пектиновых веществ, слоев радиально расположенного воска и распределенного между ними в беспорядке кутина. Самый внутренний слой оболочки, прилегающий к полости клетки, кутина не содержит. Воск может откладываться не только внутри, но и снаружи оболочки в виде мелких зернышек, тонких палочек или сплошных корочек, образуя восковой налет.
Кутин обладает как гидрофильными, так и гидрофобными группами, поэтому в кутикулярных слоях оболочки он может выполнять роль цемента, скрепляющего вместе гидрофильные полисахариды и гидрофобный воск. Кутинизация оболочки рассматривается как приспособление для уменьшения испарения (транспирации) и вымывания веществ из клеток выпадающими осадками. Кутикула защищает клетки от проникновения паразитов и механических повреждений. Она поглощает ультрафиолетовые лучи и поэтому может служить поверхностным радиационным экраном.
При ослизиении клеточных оболочек образуются слизи и близкие к ним камеди. Они представляют собой полимерные углеводы, родственные пектиновым веществам, и отличаются способностью к сильному набуханию при соприкосновении с водой. Набухание очень часто сопровождается их сильным разжижением вплоть до полного растворения. Между слизями и камедями трудно провести четкие различия. Камеди в набухшем состоянии клейки и могут вытягиваться в нити, тогда как слизи сильно расплываются и в нити не вытягиваются.
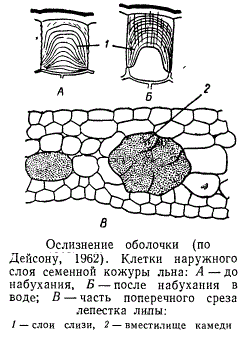
Ослизнение клеточной оболочки
Ослизнению могут подвергаться как пектиновые вещества оболочки, так и целлюлоза. В первом случае говорят о пектиновых слизях (лилейные, крестоцветные, мальвовые, липовые, розоцветные), во втором случае — о целлюлозных слизях, встречающихся значительно более редко (например, у орхидных). В семенах горчицы, некоторых бобовых и подорожника образуются смешанные, пекто-целлюлозные слизи.
Слизи образуются не только в результате химического видоизменения уже имеющихся веществ оболочки, они могут непосредственно выделяться протопластом при росте оболочки в
толщину. В качестве вторичного утолщения оболочки слизь откладывается, например, в наружных клетках семенной кожуры льна и айвы. При этом вся внутренняя поверхность наружных стенок оболочек может быть одета слизью, расположенной в виде последовательных слоев. Семена с такой кожурой сильно набухают в воде. Это имеет большое приспособительное значение при прорастании семян: попадая на влажную почву, они лучше поглощают почвенную влагу и воду атмосферных осадков. Слизь, благодаря клейкости, закрепляет семя на определенном месте, улучшает водный режим всхода и защищает его от высыхания. В состав оболочек корневых волосков входит довольно много пектиновых веществ. Ослизняясь, корневые волоски прочно склеиваются с частицами почвы. Образование слизей происходит и при поранении тканей растения, в этом случае слизи выполняют защитную функцию, закрывая рану с поверхности и предохраняя ткань от инфекции.
Иногда слизи и особенно камеди могут образоваться в результате растворения и разрушения клеточных оболочек и содержимого клеток, которое носит патологический характер. Например, у вишни часто наблюдается выделение камеди (камедетечение) из пораненных участков стволов и ветвей. Вишневый клей и представляет собой застывшую камедь. Иногда разрушение оболочек клеток и превращение их в камеди происходит не на поверхности органов растений, а внутри них. В этом случае образуются вместилища камеди внутри того или иного органа.
Оболочки стареющих клеток, в особенности расположенных на поверхности надземных органов, могут содержать не только органические, но и минеральные вещества. Эти вещества, обычно аморфные, заполняют микрокапилляры оболочки и придают ей твердость и хрупкость. Процесс инкрустации оболочки минеральными веществами носит название минерализации. В качестве инкрустирующих веществ наибольшее распространение имеют кремнезем и углекислый кальций.
Отложение кремнезема наиболее характерно для клеток кожицы и волосков хвощей, злаков и осок. У тонкостенных клеток окремнение захватывает всю оболочку, тогда как у толстостенных клеток кремний откладывается слоями, и окремневшие слои чередуются с целлюлозными слоями. Приобретаемая в результате окремнения жесткость стеблей и листьев служит защитным средством против мягкотелых животных (улиток, слизней), а также значительно снижает поедаемость и кормовую ценность этих растений. Поэтому осоки и злаки рекомендуется скашивать до цветения, после которого начинается усиленное окремнение оболочек и сено грубеет.
Углекислый кальций у высших растений откладывается главным образом в волосках, которые становятся хрупкими (например, в семействах бурачниковых, крестоцветных и тыквенных). Об отложении углекислого кальция в цистолитах говорилось выше.