Изучение цитоплазматической наследственности на современном этапе развития генетики представляет существенный интерес для выяснения генетической системы клетки в целом.
Теперь становится очевидным, что познание наследственности возможно только при условии установления взаимосвязи ядра и цитоплазмы. Это вытекает из фактического существования двух типов наследственности: хромосомной наследственности и внехромосомной (цитоплазматической) наследственности, а также соответственно хромосомных и внехромосомных генов.
Прежде чем приступить к систематизации методов генетического анализа цитоплазматической наследственности, необходимо установить терминологию.
До сих пор под термином «генотип» мы имели в виду лишь ядерную или хромосомную наследственность, т. е. систему генов, локализованных в хромосомах.
Наследственные факторы, локализованные в цитоплазме (и ее органоидах), обычно обозначаются термином «плазмотип», иногда «плазмон» или «цитоплазмон». Мы принимаем термин «плазмотип». Единицу цитоплазматической наследственности, соответствующую единице хромосомной наследственности (гену), называют плазмогеном.
Совокупность генотипа и плазмотипа, образующих генетическую систему клетки, иногда называют идиотипом. С нашей точки зрения, правильнее было бы понятие генотипа расширить и придать ему содержание термина идиотипа, а совокупность наследственных факторов ядра обозначить термином «нуклеотип» — ядерная наследственность; совокупность наследственных факторов, локализованных в цитоплазме и ее органоидах, можно обозначить термином «цитотип» или сохранить термин «плазмотип». Таким образом, термин «идиотип» будет излишним.
Расширенное понятие генотипа будет включать в себя всю систему наследственности клетки, что наилучшим образом соответствует современному представлению о наследственности.
Трудности генетического анализа цитоплазматической наследственности объясняются следующими существующими в данное время обстоятельствами:
- не известна точная локализация цитоплазматических наследственных факторов;
- не определена природа дискретной единицы цитоплазматической наследственности — внеядерного гена;
- отсутствуют статистические закономерности расщепления из-за неменделевского типа наследования;
- не установлены группы сцепления;
- не определено число факторов, наследуемых через цитоплазму, и органоидов, передаваемых при делении дочерним клеткам;
- неравный вклад цитоплазматической наследственной информации со стороны отцовского и материнского организмов в зиготу и возможность при этом материнского типа наследования.
В поисках методов проведения генетического анализа цитоплазматической наследственности, по-видимому, следует отказаться от исходного предположения, что механизмы ядерной и цитоплазматической наследственности могут быть тождественны. Возможно, что они либо разные, либо цитоплазматической наследственности как особого механизма вообще не существует, и она представляет собой лишь систему реализации действия ядерных генов с особым механизмом передачи генных продуктов через органоиды цитоплазмы или простое воспроизведение детерминированных ядерными генами структур — органоидов, обладающих способностью воспроизводиться в клетке. В этом направлении требуются дальнейшие исследования.
Однако отвергать испытанные и проверенные генетические методы анализа генотипа для изучения цитоплазматической наследственности преждевременно. Совершенно очевидно, что для генетического анализа цитоплазматической наследственности необходимы точные цитологические и биохимические данные об органоидах клетки — об их морфологии, постоянстве или закономерностях изменения их числа в клеточном цикле, об их воспроизведении и функции. Для успешного исследования цитоплазматической наследственности генетическими методами сначала необходимо исключить предетерминацию цитоплазмы, инфекцию всех типов, преадаптацию цитоплазмы и яйцеклетки во время развития ее и наследование генов, сцепленных с полом и с хромосомными перестройками. Примером этого может служить наследование сцепленных Х-хромосом (уу), нехватка типа Notch и др.
Одним из основных методов генетического анализа цитоплазматической наследственности является получение независимых мутаций, их идентификация и проверка наследования в ряду поколений.
Для исследования цитоплазматической наследственности важно получить цитоплазматические мутации, затрагивающие различные четко проявляющиеся признаки как морфологического, так и биохимического характера, связанные с различными органоидами клетки. Такие мутации могут касаться пластид, митохондрий, вероятно, рибосом, центриоли и других воспроизводящихся в митотическом цикле клетки структур цитоплазмы. Лишь при накоплении мутантов становится возможным исследовать расщепление, рекомбинации и локализацию.
Как мы уже знаем, у высших растений пестролистные мутанты в ряде случаев являются примером возникновения пластидных мутаций. Однако цитоплазматические мутации спонтанно проявляются редко, а индукция их рентгеновыми лучами оказывается мало эффективной. Б. С. Эфрусси предложил метод массового получения цитоплазматических мутаций у дрожжей путем обработки их акридиновыми красителями. Последние оказались наиболее эффективными в вызывании цитоплазматических мутаций не только у дрожжей, но и у грибов (например, у нейроспоры).
А. Срб индуцировал новые цитоплазматические мутации у нейроспоры Neurospora sitophila, используя предложенный Б. С. Эфрусси метод обработки акридиновыми красителями. Был получен ряд независимо возникших мутантов; все они отличались от дикого типа N более медленным ростом. Вначале все выделенные варианты проявляли нестабильность, однако среди них оказались штаммы с четкой характеристикой замедленного роста slow growth — Sg. Эти мутанты характеризовались замедленным прорастанием спор и конидий. После прорастания рост у этих мутантов идет с нормальной скоростью. Анализ цитохромного спектра мутантов Sg не обнаружил отличий от дикого типа, следовательно, замедленное прорастание не было связано с нарушением дыхания.
Изучение такого типа мутаций в реципрокных скрещиваниях с диким типом показало материнский тип наследования. Исследование беккроссов: FB1 (♀Sg x ♂N) x N в ряду поколений (до двадцати беккроссов FB20) показало, что при полном замещении материнских хромосом хромосомами отцовского типа признак Sg неизменно наследовался по материнской линии.
Испытание мутанта Sg на сцепление с хромосомными генами показало, что Sg не сцеплен ни с одним из генов всех 7 хромосом нейроспоры. С помощью тетрадного анализа выяснилось, что отсутствие расщепления по Sg не влияет на расщепление по хромосомным генам (4 : 4).
Таким образом, цитоплазматический характер наследования Sg не может быть объяснен какими-либо нарушениями в ядре или особенностями мейоза.
Было выделено 6 различных мутаций Sg среди 51 независимо возникшей. Каждая из 6 мутаций характеризовалась специфическим отличием в скорости роста от дикого типа. Проверка в реципрокных скрещиваниях мутантов Sg и poky: Sg x poky → Sg, poky x Sd → poky, показала, что они действительно являются различными цитоплазматическими мутантами.
Далее было произведено смешение цитоплазмы — экспериментальное слияние гифов двух форм нейроспоры: с нормальной цитоплазмой N и хромосомным маркером a (albino — белый мицелий) и Sga+. Вскоре после того, как под микроскопом наблюдалось слияние гифов, все появляющиеся в этом участке отростки с ядрами отсекали и переносили в отдельные пробирки. С выросших мицелиев собирали и анализировали конидии (споры бесполого размножения). Среди выросших колоний были обнаружены не только исходные сочетания Na и Sga+, но и новые сочетания Sga и Na+. Таким образом было обнаружено расщепление по цитоплазматическому фактору, причем оно оказалось независимым от распределения ядер.
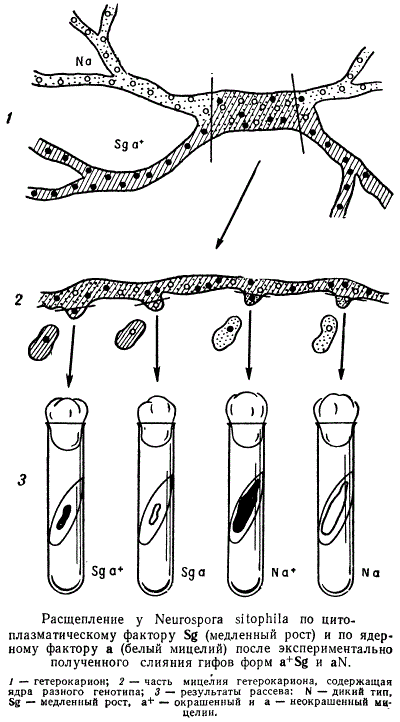
Важно обратить внимание на то, что иногда внеядерные гены могут наследоваться и по отцовской линии. Так, например, у нейроспоры был выделен мутант АС_7 с медленным, но равномерным ростом и с нарушениями в системе цитохромов. В этом случае при реципрокном скрещивании мутанта АС_7 x N и N x АС_7 все потомство оказывалось дикого типа, т. е. тип АС_7, поддерживаемый при бесполом размножении, как бы исчез при скрещивании независимо от его направления. Затем штамм АС_7 был маркирован геном а. В прямом и реципрокном скрещиваниях расщепление по этому гену наблюдалось нормальное. Следовательно, нарушений в мейозе нет. Но прямое и реципрокное скрещивания штаммов с цитоплазматическими мутациями АС_7 и Sg (AC_7 x Sg и Sg x AC_7) дают в потомстве только Sg.
Таким образом, в этом скрещивании признак Sg, передававшийся до того потомству исключительно по материнской линии, оказался переданным и через микроконидии. Механизм такой передачи цитоплазматических мутаций остается неясным. Однако приведенный пример не является исключением. Такое явление установлено также у хламидомонады.
Как мы отмечали, спонтанные цитоплазматические мутации выявляются реже, чем мутации хромосомных генов. Это можно объяснить рядом причин. Очевидно, одна из причин лежит во множественности цитоплазматических структур и органоидов. Всякая цитоплазматическая мутация, возникшая в одном из многих идентичных органоидов, не может проявиться до тех пор, пока она не размножится в цитоплазме клетки. Цитоплазматическая мутация может проявиться в двух случаях: если данный органоид в клетке является единичным или представлен малым и постоянным числом, либо если мутаген имеет специфическое действие на органоиды клетки, вызывая массовое изменение их.
Для изучения цитоплазматических мутаций очень удобным объектом оказалась хламидомонада. Р. Сэгер удалось установить, что стрептомицин вызывает у нее большое количество мутаций нехромосомных генов. При обработке раствором стрептомицина штаммов, чувствительных к этому антибиотику (Sms), были выделены мутанты с хромосомными и цитоплазматическими маркерами устойчивости к стрептомицину (Smr) и зависимости от него (Smd). Далее речь пойдет только о цитоплазматических мутациях.
Стрептомицин оказался неспецифическим сильным мутагеном в отношении разного рода цитоплазматических мутаций. При этом выяснилось, что при обработке стрептомицином частота мутаций нехромосомных генов гораздо выше, чем частота мутаций хромосомных генов. Теперь выделено несколько десятков различных цитоплазматических мутантов (около 30), фенотипический эффект которых сходен с эффектом соответствующих хромосомных мутаций. Как правило, все эти мутации наследуются по материнской линии, хотя зооспоры и материнского, и отцовского типов привносят в зиготу цитоплазму. Лишь некоторые мутанты наследуют свои признаки через обоих родителей, как это было показано для нейроспоры.
Анализ наследования цитоплазматических мутаций у хламидомонад иногда позволяет обнаруживать явление расщепления и рекомбинаций. Такое расщепление может быть равным 1 : 1 и не равным 1:1. При этом расщепление по нехромосомным генам начинается в ходе первых митотических делений после мейоза. После 4—5 таких делений 50—60% потомства все еще содержат оба родительских альтернативных нехромосомных гена, остальные другие сочетания. Рекомбинация по нехромосомным генам может приводить к возникновению как форм дикого типа, так и новых сочетаний цитоплазматических мутаций. Полученные данные свидетельствуют о том, что после мейоза каждый из таких нехромосомных генов представлен в единичном числе или в количестве лишь нескольких копий.
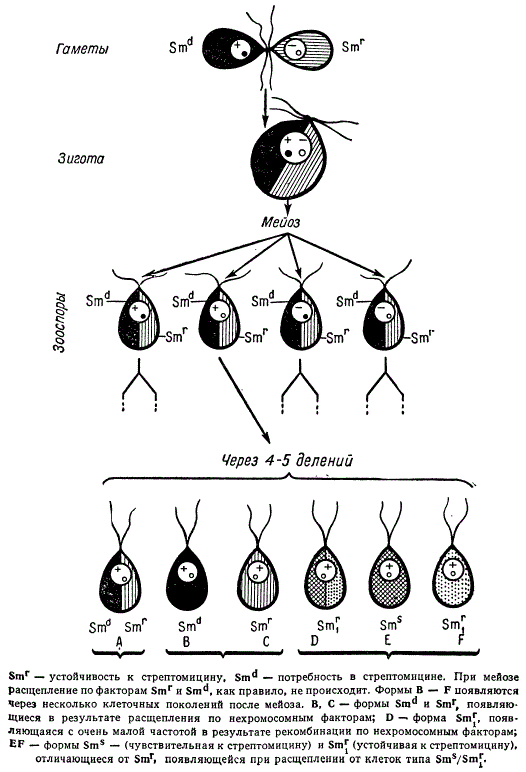
Скрещивание форм Chlamidomonas reinhardii, расщепляющихся по признакам, наследующимся через цитоплазму
Высказывается предположение о том, что может существовать упорядоченный механизм распределения цитоплазматических носителей нехромосомных генов при делении клетки. Однако локализация и форма существования таких генов остаются загадкой. Это первые шаги в изучении дискретности цитоплазматической наследственности; они могут оказаться ошибочными, но в смысле первой ориентации полезны и перспективны для раскрытия генетических тайн цитоплазмы. Эти шаги показали возможность генетического анализа цитоплазматической наследственности.
Итак, на основе изложенных фактов цитоплазматической наследственности можно сделать некоторые выводы.
1. Роль цитоплазмы в наследовании ряда признаков с несомненностью установлена у самых различных организмов. Это убедительно свидетельствует о том, что материальная и функциональная преемственность между поколениями обеспечивается всеми самовоспроизводящимися структурами клетки: ядерными и цитоплазматическими.
2. Цитоплазматическая наследственность дискретна: ее можно изучать путем спонтанных и индуцированных мутаций — плазмогенов.
3. Тщательное исследование явлений цитоплазматической наследственности будет способствовать решению многих генетических и общебиологических проблем, из которых наиболее важной является проблема взаимодействия ядра и цитоплазмы в процессе передачи наследственной информации и реализации ее в онтогенезе.