Все же гетеротопии и гетерохронии имеют ограниченное филогенетическое значение. Сами по себе они остаются все же лишь смещениями развития органов во времени и по месту закладки. Если они и приобретают большое значение, то главным образом потому, что часто связываются с коренной перестройкой органов. Например, смещение сердца у птиц и млекопитающих вглубь грудной клетки связано с четырехкамерным строением его. Однако, это изменение строения сердца только сопутствует данной гетеротопии. Самая же перестройка органа является следствием другого типа изменений, а именно — филэмбриогенезов.
«Теория филэмбриогенезов пытается разрешить вопрос о том, как происходят и в какой период индивидуальной жизни возникают те изменения, которые ведут к филогенетическому преобразованию взрослого организма» (Северцов, 1939). Из сказанного видно, что под филэмбриогенезами понимаются такие изменения в онтогенезе, которые определяют собой возможности новых направлений филогенеза.
Предварительно укажем, что филэмбриогенезы, как правило, возникают в течение периода морфогенеза (формообразование органов), следовательно, в эмбриональный период и являются эмбриогенезами. Совершенно ясно, что источником филэмбриогенезов являются наследственные изменения. Последние фенотипически выражаются в конкретном эмбриональном изменении. Если данное наследственное изменение вызвало резкое нарушение в ходе морфогенеза (в развитии морфогенетических корреляций), то оно не сможет получить значения филэмбриогенеза, так как организм вообще не достигнет взрослого состояния. Если этого не случилось, то наследственно обусловленное эмбриональное изменение войдет в онтогенез, станет его элементом к будет повторяться у потомков, влияя на строение взрослого организма. Если при этом окажется, что данное эмбриональное изменение полезно (или не вредно), и притом как во время эмбриональной, так и взрослой фазы онтогенеза, то оно станет филэмбриогенезом, т. е. таким эмбриональным изменением (эмбриогенезом), которое влияет на направление филогенетического развития потомков (филэмбриогенез).
Таким образом, филэмбриогенезы, с которыми, как правило, мы имеем дело у сложившихся видов, образовались давно и являются свидетельством когда-то возникшего эмбрионального наследственного изменения. Именно этот факт и имеет огромное значение в исследовании филогенеза, так как изучая такие филэмбриогенезы, зафиксированные в онтогенетическом развитии, мы получаем возможность судить о том, по каким направлениям и какими способами происходила эволюции данной группы.
Разумеется, мы можем быть свидетелями новых наследственные изменений, а следовательно, новых эмбриогенезов, которые являются зачинающимися филэмбриогенезами.
Выяснив вопрос об отношениях между филэмбриогенезами и наследственными изменениями, перейдем к вопросу о времени возникновения и типах филэмбриогенезов.
Было уже указано, что возникновение филэмбриогенезов падает преимущественно на период морфогенеза (Северцов), следовательно, в основном, на эмбриональную фазу индивидуального развития, когда, как мы видели, завязываются морфогенетические корреляции. Таким образом, взрослое животное вступает в жизненный путь с новыми признаками — последствием новых филэмбриогенетических изменений.
Продолжительные исследования Северцова позволили ему установить следующие основные способы филэмбриогенетических изменений.
А. Способ анаболии, или надставок конечных стадий зародышевого развития. Северцов приводит следующий пример. У саргана (Belone acus) челюсти резко удлинены, так что обе они образуют сильно вытянутое рыло. Изучая зародышей саргана, Северцов нашел, что у эмбрионов в 10 мм длины обе челюсти еще короткие, причем нижняя челюсть слегка выступает вперед. По мере дальнейшего развития, нижняя челюсть обгоняет в росте верхнюю. Затем начинается рост и верхней челюсти. Получается форма челюстного аппарата, изображенная на рисунке. На этой стадии развития малёк саргана сильно напоминает другую взрослую рыбу Hemirhamphus. В дальнейшем верхняя челюсть малька саргана продолжает расти и в результате формируется удлиненное рыло взрослого саргана.
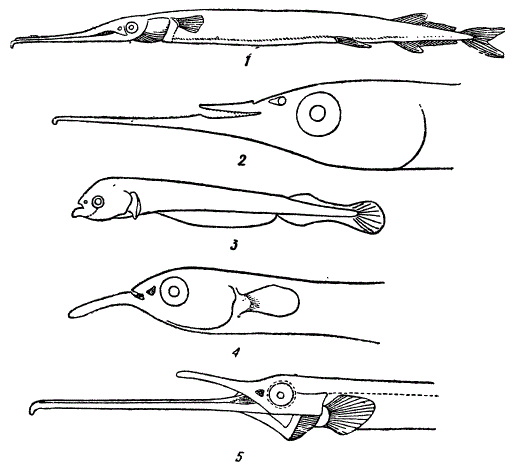
Филэмбриогенезы. 1 — взрослая особь Belone acus, 2 — голова взрослого Hemirhamphus, 3 — малек В. асus 10 мм длиной, 4 — малек В. acus 21 мм длиной, 5 — малек В. acus 9,1 см длиной (По Северцову)
Замечательно, что у других представителей того же семейства рыб (Scombresocidae) строение рыла напоминает стадию малька саргана, с еще короткими челюстями. Таким образом, у саргана формообразование челюстного аппарата изменилось по сравнению с другими представителями того же семейства; к прежним стадиям развития и роста прибавились еще две конечные стадии: «стадия Hemirhamphus’a (как ее можно условно назвать) и стадия «взрослого саргана». Произошла, следовательно, надставка стадий эмбриогенеза, или анаболия. Легко видеть, что на ранних стадиях эмбриональное развитие саргана рекапитулирует ход развития, свойственный другим рыбам того же семейства. Но на последней стадии развития наблюдается надстройка над прежними стадиями — анаболия.
Б. Мы видели, что уже Мюллер указывал на другой способ возникновения филэмбриогенезов: отклонение от типичного развития на одной из средних стадий эмбриогенеза. Этот способ Северцов называет способом девиации (аберрации).
Хороший пример девиации был предложен Матвеевым (1932). На рисунке показано развитие плакоидной чешуи акулы Heptanchus cinereus. В коже образуется комплекс клеток, который постепенно разрастается по направлению к поверхности ее, причем за счет сложных преобразований в соединительнотканном сосочке образующейся плакоидной чешуи развивается костный плакоидный зуб.
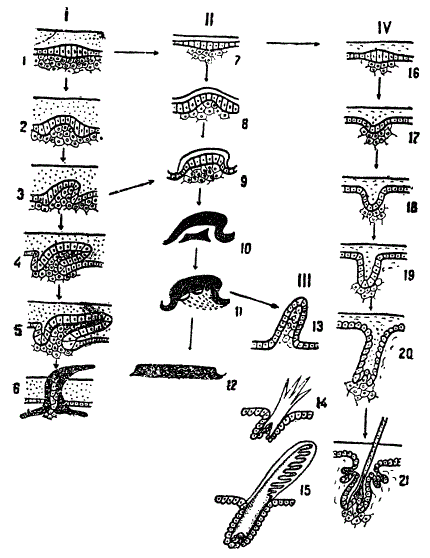
Схема филогенетического развития костных и роговых чешуй, пера и волоса.
1—6 развитие чешуи рыбы, 7—12 — чешуи пресмыкающегося, 13—15 — пера птицы, 16—21 — волоса млекопитающего. Первые стадии процесса всюду сходны. Развитие чешуи пресмыкающегося сперва рекапитулирует развитие чешуи рыб, затем происходит девиация (9). Развитие пера птиц сперва рекапитулирует развитие чешуи пресмыкающегося, затем происходит анаболия (11) и на конечных стадиях развитие пера идет специфично. Развитие волоса уже на ранней стадии (17) уклоняется от предшествовавших типов формообразования (архаллаксис). Из Матвеева, схематизировано
Исследуя развитие чешуи у стенной ящерицы Lacerta muralis, Матвеев нашел, что первоначально оно происходит по типу развития рыбьей чешуи, однако на одной из средних стадий развитие чешуи ящерицы отклоняется от этого типичного для рыб пути. Прогрессивным преобразованиям подвергается не соединительнотканный сосочек будущей чешуи, но ее эпидермальная часть, которая образует не костную, а роговую дефинитивную чешую ящерицы. Рисунок отчетливо показывает, что это отклонение произошло на средних стадиях развития. Следовательно, чешуя рептилии образуется по способу девиации. При этом способе первые стадии рекапитулируют более древний («рыбий») способ развития, ведущий к образованию костной чешуи, но на дальнейших стадиях девиация приводит к образованию роговой чешуи нового типа — «рептильного». С момента возникновения девиации повторения (рекапитуляции) уже нет. Как видим, на преемственно старой основе развивается новое направление эволюции кожных покровов.
В. Третий способ возникновения филэмбриогенезов Северцов называет apxaллаксисом, под которым подразумевается изменение, возникшее на начальных стадиях эмбрионального развития данного органа или его части. На рисунке показан интересный случай архаллаксиса в развитии волоса млекопитающего. Еще в начале развития волоса под эпидермальным комплексом клеток образуется скопление соединительнотканных клеток, за счет которых формируется луковица волоса. Таким образом, зачаток волоса состоит из комплекса эпидермальных и соединительнотканных клеток. Таков же зачаток чешуи акулы. Следовательно, закладка волоса млекопитающего гомологична закладке чешуи «рыбьего» типа. Однако, уже на первой стадии развития волоса прогрессивное развитие претерпевает не соединительнотканный комплекс клеток (как у рыб), а эпидермальный, который энергично растет, врастает в собственно кожу и превращается в стержень волоса, одетый волосяной сумкой. Таким образом, волос начинает свое развитие как чешуя, но на первых же шагах направление его развития изменяется и идет по новому пути, ведущему к образованию волоса. Очевидно, что формирование волоса достигается по способу архаллаксиса, при котором рекапитуляция исключена полностью.
Указанные три способа филэмбриогенетических изменений могут быть для наглядности выражены следующими буквенными символами:
а) способ анаболии
предок: a, b, с, d, D
потомки: a, b, с, d, е, E; а, b, с, d, е, f, F,
где a, b, с, d и т. д. обозначают эмбриональные стадии, а большие буквы — взрослое состояние.
б) способ девиации:
предок: а, b, с, d, е, f, F, потомок: а, b, с, d, с1, f1, F1.
в) способ архаллаксиса:
предок: а, b, с; d, е, f, F, потомок: a1, b1, с1, dl, е1, f1, F1
Бросается в глаза, что при архаллаксисе рекапитуляция исключена с первой же стадии (а1, b1, с1 и т. д.).
Все три способа филэмбриогенетических изменений наблюдаются в системе одного и того же организма. Однако, анаболия наблюдается значительно чаще, как это было подмечено уже Дарвином, который придавал наибольшее эволюционное значение изменениям, возникшим на более поздних стадиях развития. Причины, по которым именно анаболии имеют наибольшее значение, вполне понятны. Изменения, возникшие на ранних стадиях (архаллаксисы), влекут за собою коренные изменения в последующем формообразовании и, следовательно, могут вызвать такие перестройки в органах, которые сделают организм нежизнеспособным. Анаболии возникают тогда, когда процесс морфогенеза, в основном, закончен, и, следовательно, резких нарушений его нет. Поэтому архаллаксисы наблюдаются, как правило, в более автономных системах, изменения в которых (например, в коже) не влияют на общий эмбриональный морфогенез (Шмальгаузен, 1942).
Так как разные органы изменяются разными способами, то мы можем прослеживать не только эволюцию организма, как целого, но и его отдельных органов.