Некоторые изменения фенотипа, обусловленные мутациями или комбинацией генов в генотипе, могут копироваться ненаследственными изменениями.
Такие копии мутантов называются фенокопиями. Механизм их появления еще не изучен. Одни из них возникают под влиянием пока еще невыясненных условий жизни, другие удалось получать экстремальным воздействием. Такие изменения, вызванные экспериментально, называют морфозами.
Причинами появления фенокопий и морфозов являются, по-видимому, изменения в цепи формообразовательных процессов.
Организм в ходе индивидуального развития представляет собой целостную, неделимую рабочую систему. В онтогенезе нельзя изменить какую-либо одну функцию (реакцию) и элементарную структуру без того, чтобы при этом не затронуть всю систему, которая складывается из последовательных, строго регламентированных морфо-физиологических процессов.
Но, признавая целостность онтогенеза, нельзя забывать о существовании отчетливой прерывности — дискретности этого процесса, выражающейся в морфогенезе и становлении функций. Биологи давно пришли к выводу, что онтогенез протекает неравномерно, что в ходе его происходит качественная смена процессов, проявляющаяся в изменении характера роста и дифференцировки.
Без признания прерывности, или, точнее, неравномерности развития нельзя было бы понять механизм приспособительных ответных реакций организма на воздействие различных факторов внешней среды,, эволюционное усложнение онтогенеза и его дискретную генетическую детерминацию. Подобную закономерность в развитии животных называют фазностью, а в развитии растений — стадийностью.
Дискретность онтогенеза была выявлена на основе изучения действия внешних факторов на индивидуальное развитие. И. В. Мичурин одним из первых обратил внимание на то, что растения, особенно многолетние, проходят различные этапы, стадии развития. Он использовал принцип дискретности для разработки метода воспитания гибридов в индивидуальном развитии. Для нормального развития растения в определенные этапы необходимы адекватные экологические факторы, что также определяется генотипом.
У растений известны озимые и яровые формы. При скрещивании гомозиготных озимых и яровых пшениц в первом поколении доминирует свойство яровости.
Во втором поколении наблюдается расщепление на яровые и озимые. Существуют также особые генотипически определяемые формы, так называемые «двуручки», которые обладают обоими свойствами — и яровостью, и озимостью, проявляющимися в зависимости от условий выращивания. Т. Д. Лысенко в 20-х годах показал, что эти свойства в онтогенезе можно изменять. Обычно озимые растения (пшеница, рожь), посеянные весной, при высоких температурах растут, кустятся, но не развиваются — колошения не наступает. Если же семена перед посевом в течение определенного времени подвергнуть действию пониженных температур при определенной влажности (яровизация), а затем высеять весной в поле, то они нормально разовьются и выколосятся.
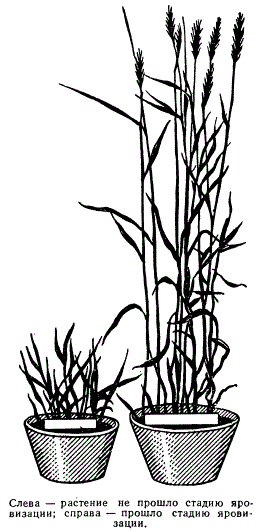
Характер развития озимой пшеницы в зависимости от прохождения стадии яровизации
Эта стадия получила название стадии яровизации. Эти изменения не наследуются, но они имитируют фенотип наследственно яровых форм, т. е. являются фенокопиями.
В связи с этими ценными в практическом отношении опытами были сделаны далеко идущие и неправильные выводы о том, что если воздействовать холодом на яровое растение в соответствующий момент развития, то в его потомстве можно получить наследственно озимые формы. Таким образом, на основе ошибочной трактовки опытов с так называемой переделкой наследственной природы растения постулировалось принципиально ошибочное положение о возможности направленного наследственного изменения признаков путем «воспитания». Заметим, что еще в 1929 г., когда впервые Р. Гольдшмидт и В. Иоллос обратили внимание на фенокопии, вызываемые высокой температурой у дрозофилы, то на первых порах они также пришли к ложным выводам о направленном превращении фенокопий в мутации, но после критической проверки опытов быстро от них отказались.
Наследственное изменение свойства озимости и яровости действительно возможно с помощью прямого влияния температурного фактора, но для этого известен пока один путь — отбор растений с генотипом, отзывчивым на влияние температуры. Хотя генетика озимости и яровости исследована слабо, однако имеются указания на то, что эти свойства определяются полигенно. Так, уже установлены по крайней мере три гена с сериями аллелей, которые детерминируют данное свойство. В силу сложности наследования при расщеплении в потомстве получаются все переходы от озимости к яровости. Промежуточные формы — «двуручки» также определяются сочетанием этих аллелей.
Выше мы уже приводили подобные примеры с реакцией некоторых летальных мутантов на температуру. Очевидно, что свойства озимости и яровости как сложные физиологические признаки определяются генотипами, которые подвергаются отбору.
100 лет назад, иллюстрируя значение отбора для превращения озимых сортов в яровые и обратно, Ч. Дарвин писал: «Пшеница быстро приспособляется к новому образу жизни. Линней отнес яровые и озимые сорта к отдельным видам; но Монье доказал, что различие между ними лишь временное. Он посеял озимую пшеницу весной, и из сотен растений только четыре принесли зрелые семена; эти семена он посеял и потом вновь сделал такой же посев: через три года получились растения, на которых все семена вызревали. Наоборот, почти все растения, выращенные из яровой пшеницы, при посеве осенью погибли от мороза, но несколько экземпляров уцелело и принесло семена; через три года эта яровая разновидность превратилась в озимую».
Дарвин, а с ним и все дарвинисты рассматривали такие опыты как яркое доказательство могущества искусственного отбора генотипов. Путем отбора на провокационном фоне (посев озимых весной, а яровых под зиму, и т. д.) создается иллюзия наследования приобретенных признаков, так как факторы внешней среды одновременно и вызывают адаптивные фенокопии, и отбирают соответствующие генотипы. Данный тип отбора был назван И. И. Шмальгаузеном стабилизирующим отбором. С нашей точки зрения, совпадение адаптивных фенокопий с мутантами, случайно оказывающимися адекватными, следует называть отбором на провокационном фоне — дифференцирующем, или селективном, фоне.
Фенокопии у растений могут быть вызваны также и световым фактором. Каждый вид растения для продолжения своего нормального развития на определенном его этапе нуждается в определенной длительности светового дня (световая стадия).
Исследованиями ряда авторов — Т. Д. Лысенко, А. А. Авакяна, В. И. Разумова, В. Н. Столетова и др. — было показано, что у разных видов растений продолжительность стадий развития, в частности стадий яровизации и световой, и конкретные условия, необходимые для прохождения этих стадий, зависят от происхождения растений, от их генотипа.
В начале нашего столетия было установлено, что такие же наиболее «чувствительные» к действию внешних факторов периоды имеют место и в онтогенезе животных. Их называют критическими периодами развития. В опытах с насекомыми А. С. Данилевским и др. была показана возможность изменять циклы развития, изменяя длину светового дня, влияющего на длительность диапаузы у насекомых. В течение онтогенеза критических периодов может быть несколько; как правило, они приходятся на время эмбриональной дифференцировки или усиленного морфогенеза.
Действие внешних факторов во время критических периодов в онтогенезе вызывает наибольшую изменчивость именно тех признаков, морфологическая дифференцировка которых приходится на этот период. Степень изменчивости развития признака в критический период определяется повреждающей способностью агента и генотипом организма. У разных организмов в результате повреждающего воздействия на один и тот же критический период онтогенеза возникают, как правило, однотипные морфозы. Например, воздействие любым повреждающим агентом, тормозящим развитие переднего мозгового пузыря зародыша, вызывает аномалию — циклопию у животных и человека. Даже слабые воздействия внешних факторов в момент критических периодов могут вызывать определенного типа морфозы.
Морфозы характеризуются тем, что они могут возникать массово, если действие агента приходится как раз на время критического периода, наступающего синхронно у большей части подвергавшихся воздействию особей. Так, при облучении рентгеновыми лучами и дополнительном температурном воздействии на личинок дрозофилы в критический период удается получать до 100% особей с однотипными морфозами (рентгеноморфозы), имитирующими мутантные признаки, например растопыренные крылья, вырезку на крыле и др. Подобные изменения можно вызывать и при воздействии другими агентами, в частности высокой температурой, изменением режима питания личинок. Это показывает, что действие различных агентов во время критических периодов может быть неспецифичным.
Однако действие некоторых агентов, по-видимому, может быть специфическим. Так, И. А. Рапопорт получал у дрозофилы под влиянием солей серебра высокий процент особей с желтым цветом тела, напоминающим мутантный признак yellow.
Фенокопии и морфозы, имитируя наследственные изменения, сами не наследуются, так как они представляют изменения в соматических клетках и возникают не в силу изменения самих генов, а в результате нарушения их действия, но предрасположение к возникновению определенного типа фенокопий обусловливается генотипом.
Изучение фенокопий имеет важное значение как для понимания действия гена, так и для исследования механизмов эволюции. Возникновение фенокопий под влиянием внешних агентов может указывать на то, что мутантные гены вызывают блокирование или изменение тех же процессов, на которые влияет агент, и в те же периоды развития, в которые удается получать фенокопии. Таким образом, получение фенокопий служит одним из методов изучения действия гена. Известны случаи, когда факторы внешней среды вызывают фенокопии, которые имеют приспособительное значение для организма; например, в периоды низких температур в природе появляются меланистические формы у насекомых, формы с антоциановой окраской у растений и т. д.
Дискретность онтогенеза наблюдается не только в отношении морфологических, но и в отношении физиологических процессов, сложных врожденных рефлексов. Установлено, что в онтогенезе животных двигательные рефлексы, например чесательный и лизательный у млекопитающих, клевательный и рефлекс затаивания у птиц и другие, также имеют фазность в становлении. То же самое относится к формированию сложных актов поведения животных.
Морфологическая и функциональная дискретность онтогенеза облегчает исследователям поиски путей и методов управления онтогенезом.