Дарвин в «Происхождении видов» справедливо подчеркнул, что недостаточно убедиться в самом факте изменяемости видов. Надо обратить внимание еще на одну черту этого процесса — приспособительный характер его. Эволюция осуществляется через приспособление видов к различным условиям существования.
а. В этом убеждает, прежде всего, тот факт, что при внимательном анализе любой живой формы мы обнаруживаем, что ее морфофизиологическая организация буквально пронизана адаптациями к конкретным условиям среды.
Каждый вид рассматривается экологом, как жизненная форма. Под последней следует понимать определенный тип морфофизиологической организации, соответствующий или приспособленный к определенному типу местообитания и притом ко всем ведущим, главным особенностям его.
Именно эта характерная черта жизненной формы с точки зрения эволюционной теории Дарвина особенно интересна и важна. Вид, как жизненная форма, оказывается (как правило) приспособленным к свойственной ему обстановке не в одном-двух признаках, а по всем основным, жизненно важным признакам организации. Чтобы в этом убедиться, рассмотрим в качестве примера жизненную форму одного из представителей отряда насекомоядных — русской выхухоли Desmana moschata.
1. Местообитание выхухоли. Выхухоль обитает в пресноводных водоемах— затонах рек и поемных озерах. Животное теснейшим образом связано с водной средой. Норы выхухоль делает в берегах, причем единственное отверстие норы расположено ниже уровня воды. Следовательно, животное нормально из норы может попасть только под воду. Питается выхухоль мелкими водяными животными (пиявки, водяные насекомые, лягушки, рыбы) и водными растениями. Водная среда служит и кормовой базой выхухоли.
Открытой суши животное, как правило, избегает. Водная среда речных затонов и поемных озер — нормальная для выхухоли среда жизни. В пределах последней выхухоль выполняет все свои жизненные функции: питание, индивидуальное развитие и размножение.
2. Образ жизни выхухоли и адаптации к нему. Подводная кормовая база выхухоли обильна. Однако, как и все насекомоядные (Insectivora), выхухоль нуждается в частом питании. Поэтому пребывание в норе, как правило, кратковременно, и значительную часть суток выхухоль рыщет по дну водоема (обычно по определенным путям) в поисках пищи, которую она либо заглатывает под водой, например, пиявок, либо поедает в надводной части особых кормовых нор (например, прудовиков).
Поиски пищи, по преимуществу донного и придонного происхождения, удерживают выхухоль под водой, что выгодно также как естественное укрытие от возможных воздушных врагов. С другой стороны, легочное дыхание диктует необходимость подниматься к поверхности, оставаясь при этом, по возможности, в скрытом состоянии и производя передвижения вверх и вниз с максимальной быстротой. Осуществление всех этих условий требует специальных приспособлений.
Рассмотрим приспособления ее к нырянию и подводному плаванию. Главным органом, обеспечивающим ныряние и плавание, является длинный ланцетовидный хвост. Он сильно сжат с боков и почти лишен волосяного покрова. Ткани хвоста очень богаты жировыми отложениями, вследствие чего удельный вес его невелик, — обстоятельство, увеличивающее его подвижность в воде. Поэтому, при движениях выхухоли по земле, хвост волочится и оставляет явственную черту, в воде же он силен и подвижен. В основании хвоста расположены мускусные железы. Их жирные выделения до известной степени смазывают весь хвост. Эта обмазка усиливает динамику, уменьшая трение о воду. Хвост выхухоли работает в горизонтальной плоскости, как орган руления.
Дополнительным органом плаванья служат попеременно гребущие задние лапы, вооруженные большими, достигающими когтей, плавательными перепонками. Подводное плаванье облегчается также хорошей обтекаемостью тела и несмачиваемостью меха. Ушные раковины выхухоли, если можно так выразиться, погружены в слуховой проход и, следовательно, не мешают движению. Носовые отверстия запираются в воде клапанами.
Движение выхухоли вверх производится автоматически, подъемной силой воздуха, заключенного в волосяном покрове. Насыщенность последнего воздухом обусловлена пышным развитием подпуши — главного резервуара воздуха, а также легкой полегаемостью остевых волос, имеющих своеобразную форму, облегчающую плотное прилегание остей к меховому слою во время подводных экскурсий. Воздуха в мехе так много, что выхухоль поднимается автоматически. Прекратив работу конечностей и приподняв кверху хоботок, она выбрасывается на поверхность с быстротой пробки. Свойства меха позволяют выхухоли свободно, подобно поплавку, лежать на поверхности, лишь частично погрузившись в воду.

Выхухоль (Desmana moschata). Вверху самец и самка под водой в момент «игры». Ниже — при плавании на поверхности воды. Рядом на земле, в позе ориентировки в пространстве. (Ориг. рис. В. В. Ватагина)
Лежа на воде, животное прожевывает пищу, или, перевалившись на бок, прочесывает задней лапой шерсть, обычно от хвоста к голове. Поражает свобода и непринужденность, с которой выхухоль ворочается на воде, как на мягкой подушке. Ныряние же осуществляется путем преодоления подъемной силы воздушно-мехового покрова.
Это явление свойственно и другим водным млекопитающим. Молодые бобрята, например, не способны нырять, так как не могут преодолеть подъемной силы своего воздушно-мехового покрова (Федюшин, 1935). Этим чертам приспособительной организации соответствует приспособительное поведение. Лежа на воде, выхухоль находится в позе «готовности к нырянию». Задние лапы поставлены так, что их гребущие поверхности ориентированы по бокам тела в перпендикулярной плоскости. Обладая плохим зрением, выхухоль хорошо слышит. Ушная раковина, как сказано выше, погружена в слуховой проход, но ее резонаторы хорошо развиты. Малейший беспокоящий звук, — тотчас раздается плещущий удар хвоста, задние ноги из исходного положения мгновенно переводятся в гребущее — и животное исчезает под водой. Подъемная сила воздушно-мехового слоя инстинктивно оберегается животным. Выйдя из воды, выхухоль тщательно сушит и пушит мех ловкими, быстрыми треплющими движениями задних ног. Распушив мех, набив в него воздух, выхухоль готова к новой водной экскурсии.
Описанный комплекс приспособлений к водной жизни пронизывает, таким образом, всю организацию животного. Легко показать, что эти приспособления, при данной организации выхухоли, в равной мере необходимы. Бесхвостые особи, например, теряют ловкость движений, в особенности при погружении. Удалось наблюдать случай, когда самка выхухоли быстро утомилась при плаванье (в аквариуме) вследствие того, что ее мех сильно вылинял и лишился воздушно-подъемных свойств. Усиленно работая хвостом и ногами, великолепно приспособленными к плаванию, животное все же целиком, до кончика носа, погрузилось в воду, начало задыхаться и тонуть. Хорошо приспособленного к плаванью зверя пришлось спасать от гибели в его родной стихии! Из этого факта можно видеть, что адаптивное значение имеет не один признак, а целая система адаптаций.
Следовательно, вся организация выхухоли, как целое, приспособлена к свойственному ей образу жизни. Ту же черту мы ранее констатировали для дельфина, и она свойственна любому виду. Таким образом, прежде всего, мы констатируем факт, что ведущей чертой организации любого вида является его всесторонняя приспособленность к конкретным условиям среды. Уже эти факты непонятны вне эволюционного учения, так как с очевидностью могут быть объяснены только как результат исторического накопления приспособлений. Без такого предположения пришлось бы допустить чудесное внезапное возникновение гармоничной организации.
б. Наше рассуждение остается, однако, лишь косвенным доказательством приспособительного характера эволюции, тогда как имеются и прямые доказательства этого. Присматриваясь, например, к эволюции лошадиных, нетрудно констатировать, что она была связана с переходом от жизни в лесах на топкой почве (эогиппус) к существованию в открытых степных просторах. Изменения образа жизни было причинно связано с изменением приспособлений. В частности от эогиппуса к современной лошади происходило:
а) Увеличение роста — важное приобретение, сопряженное прежде всего с увеличением силы животного. Кроме того — и это главное — высокий рост лошади (по сравнению с эогиппусом) является несомненным приспособлением к более дальнему видению, к расширению горизонта в условиях степных просторов.
б) Увеличение быстроты бега, достигнутое путем облегчения скелета ноги, т. е. уменьшения числа пальцев. Быстрый бег полезен и необходим в открытых пространствах, имея важное защитное значение, он обеспечивает также быстрое достижение водопоев, кормовых угодий и т. п.
в) Интенсификация перетирающей функции зубного аппарата в результате развития гребней на коренных зубах и моляризации остальных зубов — процесс очень важный в связи с использованием, в качестве корма, жесткой злаковой растительности.
Параллельно с этими процессами приспособительной эволюции происходили изменения, коррелятивно связанные с ними. Так, удлинились и изменились по форме челюсти, увеличился череп, спина приобрела вогнутый профиль и т. п. Как видим, эволюция лошадиных происходила через приспособление этой группы к степным условиям существования.
в. То, что выше показано для эволюции лошадей, можно подтвердить и для ныне живущих форм. Как правило, виды, входящие в состав одной естественной группы, захватывают в пределах общего своего ареала (например, ареала семейства) различные местообитания, вследствие чего в пределах одной естественной группы наблюдается большое многообразие жизненных форм. Так, в пределах ряда естественных групп животных мы найдем представителей следующих основных классов жизненных форм:
1-й класс — гидробионты (жители воды); 2-й класс — амфибионты (водно-наземные формы); 3-й класс — хтонобионты (наземно-живущие формы); 4-й класс — эдафобионты (землерои, жители почвы); 5-й класс — троглобионты (жители пещер); 6-й класс — дендробионты (древесные формы); 7-й класс — авиабионты (летающие формы); 8-й класс — паразиты и т. д.
Например, в пределах отряда грызунов встречаются амфибионты (ондатра, бобр), эдафобионты (слепыш, слепушонка и т. д,), троглобионты (пещерная крыса), хтонобионты (большинство грызунов), дендробионты (белка, белки-летяги — последние с чертами авиабионтов). Подобные же формы могут быть установлены для насекомоядных Insectivora, для хищников Carnivora, для различных отрядов насекомых и т. п.
Рисунок показывает, что представители одной естественной группы, в данном случае — грызуны, принадлежащие к разным жизненным формам, резко отличны по своим приспособительным признакам, в соответствии с различиями их местообитаний. Но в то же время все они сходны по систематическим признакам. Так, грызуны, весьма различные как жизненные формы, весьма сходны по строению зубного аппарата — основного диагностического признака отряда грызунов. Таким образом, на фоне единства их организации развертывается картина их приспособительного многообразия.
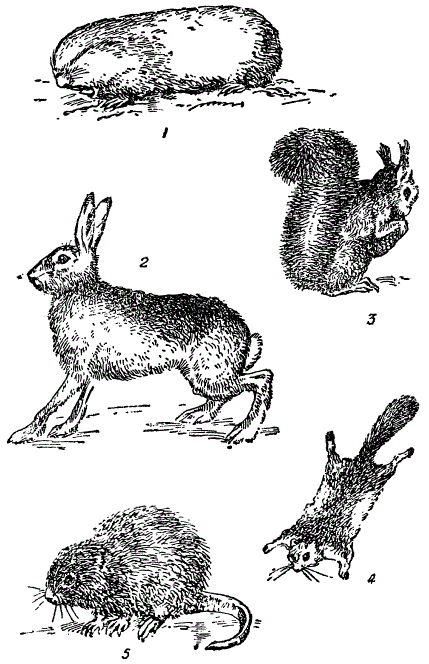
Приспособительные типы грызунов. 1 — слепыш (здафобионт), 2 — заяц-русак (хтонобионт), 3 — белка (дендробионт), 4 — белка-летяга (планирующий дендробионт), 5 — ондатра (гидробионт). Преимущественно из Бобринского
Отсюда — единственный естественный вывод, а именно, что виды одной естественной, т. е. одной монофилетической группы разошлись по своим адаптивным признакам, а следовательно, эволюировали в различных направлениях. Таким образом, приспособительная диференцировка естественной группы явственно следует закону дивергенции. Явление это носит всеобщий характер и описанную картину мы наблюдаем в пределах любой естественной группы. При этом подтверждается дарвиновская интерпретация биологического значения этого расхождения. Оно связано с диференцировкой жизненных потребностей. Приспособление к различным местообитаниям, использование максимально различной пищи позволяет видам одной естественной группы выйти из условий острой взаимной конкуренции, чем обеспечивается возможность численного роста видового состава группы. Осуществляется, следовательно, закон Дарвина о наибольшей сумме жизни при наибольшем разнообразии. Поэтому те естественные группы, которые отличаются разносторонним адаптивным расхождением, обычно богаты в видовом отношении. Среди насекомых в этом смысле выделяются жуки. Отличаясь очень полным и разносторонним адаптивным расхождением (среди них мы найдем представителей всех перечисленных выше классов жизненных форм), они в то же время очень богаты видами. Общее число видов жуков превышает 200 000. Несомненно, столь большое число видов — прямой результат не столько твердых элитр (хорошо защищающих жуков), как это обычно принято считать, сколько многообразной дивергенции. Огромное многообразие адаптаций в пределах отряда жуков прямо пропорционально их сумме жизни, и в частности, числу видов.
Справедливость сказанного хорошо демонстрируется также на других группах. Очень демонстративно, например, дивергентное развитие в пределах дневных хищных птиц. Все они плотоядны. Однако, питаясь различной животной пищей, дневные хищники диференцировались по потребностям. Так, среди дневных хищных птиц было предложено (Жарков и Теплицкий, Формозов) различать: 1) мюзифагов (мышеяды — пустельга), 2) орнитофагов (птицеяды — ястреб-тетеревятник, ястреб-перепелятник, благородные соколы), 3) рептилиефагов (орел-змееяд), 4) энтомофагов (насекомоядные — кобчик, осоед) и т. п.
Все эти хищники, имея разные потребности, различаются в значительной степени по местообитаниям, по поведению (дивергенция повадок), по типу полета, по морфологической организации, в особенности по строению крыла (дивергенция морфологической организации) и т. д.
Следовательно, внутри естественной группы осуществляется глубокая, разно направленная диференцировка организации, связанная с соответствующей диференцировкой местообитания и способов питания. Отсюда богатство видов — наибольшая сумма жизни при наибольшем разнообразии.
Итак, дивергентная эволюция форм есть наиболее общая черта исторического их преобразования.
г. В предшествующем изложении мы исходили из понятия естественной группы, дивергировавшей в различных приспособительных направлениях. Дарвин указывал, что в ходе дивергентного развития представители различных естественных групп сплошь и рядом встречаются в пределах одной и той же стации. Различные естественные группы дают вследствие этого аналогичные жизненные формы.
В этих условиях у неродственных форм, населяющих одну и ту же стацию и, следовательно, обитающих в сходных условиях и ведущих аналогичный образ жизни, развиваются сходные приспособления. Тогда систематически далекие формы, например, акула и дельфин, приобретают поверхностное сходство. Это явление сходства между неродственными жизненными формами в условиях сходного образа жизни называется конвергенцией.
Конвергенция распространена очень широко. Объясняется это тем, что стаций в природе значительно меньше, чем естественных групп организмов, почему и дивергенция многих естественных групп протекает аналогично, и они дают сходные жизненные формы. Как правило, любая естественная группа дивергирует во всех возможных (для нее) направлениях. Отсюда понятно, что в одной стации неизбежно встречаются представители самых различных естественных групп (в водной среде, например, обитают наряду с рыбами: насекомые, многие амфибии, некоторые рептилии, большое число птиц, ряд млекопитающих). Следовательно, широкое распространение конвергентного сходства между неродственными формами есть прямое и неизбежное следствие широкого распространения дивергентного развития большинства естественных групп и неизбежной встречи их представителей в пределах одной и той же экологической обстановки.
Рассмотрим некоторые примеры конвергенции, подобранные по классам жизненных форм.
1. Гидробионты дают очень много примеров конвергенции. Классический пример — конвергентное сходство между ихтиозавром, акулой и дельфином. На рисунке показано конвергентное сходство между медузой и одноклеточным жгутиконосцем Medusochloris phiale. Плазматический зонт этого простейшего жгутиконосца живо напоминает колокол медузы. Это сходство тем интереснее, что М. phiale и плавает, как медуза, т. е. при помощи ритмичного сокращения зонтика. У другого представителя жгутиконосцев — Craspedotella по краю зонтика развита оторочка, необычайно сходная с парусом (velum) гидроидных медуз.
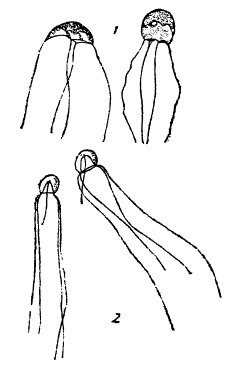
Явления конвергенции у гидробионтов. 1 — жгутиконосец Medusochloris phiale, плавающий при помощи плазматического зонта, 2 — медуза Sarsia (Coryne) tubulosa. (1 — по Патеру, 2 — из Броха)
Замечательное сходство этих форм с медузой носит конвергентный характер и не связано с родством.
Очень полный ряд конвергентных сходств дают прикрепленные, или седентарные, донные гидробионты.
У сидячих простейших, губок, кишечнополостных, кольчецов, мшанок, ракообразных, иглокожих, наконец, первичнохордовых, например, асцидий наблюдается развитие корнеподобных ризоидов, при помощи которых животные укрепляются в грунте; далее, многим из них свойственна стебельчатая форма тела — стебелек несколько компенсирует седентарность, позволяя отклоняться под ударом волны, плавника рыбы и т. п., смягчая этим силу толчка; далее, у седентарных форм, независимо от их систематического положения, мощного развития достигают защитные покровы («домики», раковины, плотные оболочки и т. д.); им свойственно также мощное развитие ресничных аппаратов, создающих ток воды и привлекающих вместе с ней пищу. Всем седентарным формам свойственна склонность к образованию колоний или же скоплений самостоятельных особей; совокупность особей, а тем более колониалыюсть, где индивидуум подчинен новому целому (колонии), снижает вероятность гибели в результате механического давления или удара. Многочисленным седентарным формам свойственна высокая регенеративная способность, снижающая и даже нивелирующая последствия ранений и т. п. (Lang, 1888).
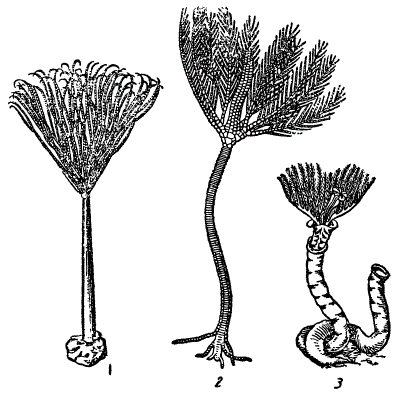
Конвергенция у седентарных (сидяче прикрепленных) животных. 1 — мшанка, 2 — морская лилия, 3 — полихета Serpula. (1,2 — из Абеля)
2. Эдафобионты. У землероев также наблюдается конвергентное сходство, яричем можно наметить ряд подчиненных групп жизненных форм, из числа которых мы коснемся только землероев типа крота. Замечательно сходство между сумчатым кротом Notorhyctes typhlops (подкласс сумчатых, сем. Dasyuridae) и златокротом (подкласс плацентарных, отряд Insectivora сем. Chrysochloridae).
Обе эти формы — песчаные землерои. Филогенетически они весьма далеки друг от друга, но поразительно сходны в ряде деталей адаптивных признаков. Так, они характеризуются совершенно сходной формой тела, наличием однородных носовых щитков, одинаковым положением наружных носовых отверстий, заросшими глазами, скрытыми в меху ушными отверстиями, очень сходным строением коренных зубов и передних конечностей и т. д.

Конвергенция у эдафобионтов. 1 — златокрот (Chrysochlons aurea), 2 — сумчатый крот (Sotoryctes typhlops. Из Северцева и Брэма
Златокроты, как и сумчатый крот, роются в песке, пользуются той же пищей. В конечном счете обе формы, ведя сходный образ жизни в идентичных стациях, приобрели совершенно изумительное сходство — настолько значительное, что было даже высказано предположение, что дело идет о кровном родстве этих форм. Однако, в действительности, сумчатый крот — типичное сумчатое, а златокрот — плацентарное млекопитающее, и, следовательно, их поразительное сходство — лишь особенно яркий случай конвергенции.
3. Дендробионты дают очень много интересных примеров конвергенции.
Так, очень многим дендробионтам свойственна хватательная конечность с оппонирующими (противопоставленными) пальцами, образующими как бы клещи для охватывания ветвей. У многих форм развитие такой хватательной конечности связано со слиянием пальцев (синдактилия). Дендробионты с описываемым типом конечностей присущи самым различным систематическим группам — от амфибий до млекопитающих включительно. Сходство хватательных конечностей у представителей указанных групп носит конвергентный характер.
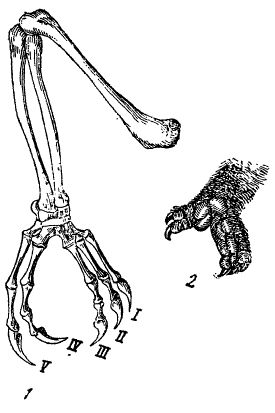
Конвергентное сходство конечностей дендробионтов. 1 — лапа хамелеона, 2 — рука кус-куса (Pseudochirus eregrinus) (по Абелю)
Общеизвестно также конвергентное сходство между планирующими дендробионтами, например, белкой-летягой Sciuropterus russicus (отряд грызунов) и сумчатыми летягами (Petauroides, Petaurus, Acrobates). Принадлежа к глубоко различным систематическим группам, не связанные пространственно, эти формы обладают изумительным сходством приспособлений к планированию.
4. Паразиты. У паразитов, как наружных (эктопаразиты), так и внутренних (эндопаразиты), также наблюдается конвергенция. Сходство между ними сплошь и рядом необычайно ярко подтверждает факт эволюции, так как филогенетически далекие формы приобретают удивительное сходство.
Так, например, паразитическая немертина Malacobdella grossa, вооруженная большой задней присоской, внешне похожа на пиявок (Hirudinea), хотя филогенетически немертины далеки от пиявок. Формы, поселяющиеся среди перьев птиц и шерсти млекопитающих, независимо от систематического положения, приобретают поразительное сходство. Так, на рисунке показаны следующие формы: вошь (отряд Anoplura) и, наконец, паразитирующая в шерсти летучих мышей муха Penicillidia noterreri (отряд Diptera). Сходство между этими формами чисто конвергентное.
Не менее эффектны случаи конвергенции среди эндопаразитов. Так, например, у многих из них имеются крючья для прикреплении к тканям хозяина. При этом форма крючьев поразительно однотипна. На рисунке показаны крючья лентецов Cestoidea и пятиусток Linguatulina.
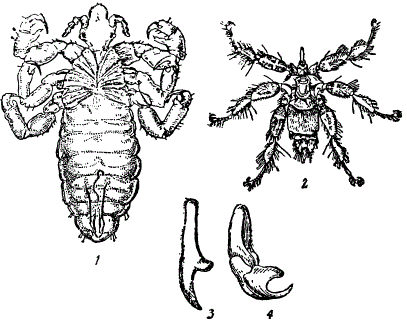
Конвергенция у паразитов. 1 — платяная вошь, 2 — муха Penicillia noterreri, паразит летучих мышей, 3 — крючок цестоды (Taenia pisiformis), 4 — крючок пятиустки Porocephalus riatae. (По разным авторам)
Их сходство весьма велико, хотя лентецы входят в состав плоских червей, а пятиустки относятся к паукообразным. Форма тела пятиусток также сходна с формой тела цестод, и долгое время их относили к червям, пока не было выяснено их истинное положение в системе.
Оставляя в стороне ряд прочих групп жизненных форм, полезно остановиться на другой стороне их классификации. В пределах любого класса жизненных форм наблюдается диференцировка его на растительноядные формы (фитофаги), плотоядные (зоофаги) и разноядные (гетерофаги). В пределах самых различных систематических групп весьма обычны все эти три группы, причем и в этом случае констатируется поразительное конвергентное сходство, в особенности в строении зубной системы.
Интереснейшим примером конвергенции нужно считать сходство зубной системы мадагаскарского лемура Chiromys madagascariensis (ай-ай) с зубной системой грызунов. Резцы ай-ай построены совершенно так же, как у последних, глубоко проникают в челюсти и постоянно растут. При этом они, как и у грызунов, покрыты эмалью только спереди, сохраняя, таким образом, острый режущий край в результате более быстрого стирания задней стенки зуба. Это сходство столь велико, что в 1-й половине XIX в. ай-ай рассматривали как представителя грызунов. Например, в сводке млекопитающих Шинца (Schinz. Synopsis mammalium 1844) ай-ай выделяется в особое семейство грызунов — Pedimana, или руконогих. В сводке млекопитающих Гибеля (Giebel, 1855) ай-ай все еще среди грызунов, хотя Гибель и сомневается, не следовало ли бы поместить это животное среди полуобезьян.
Приведенная историческая справка показывает, как велико сходство зубной системы этого лемура с зубным рядом грызунов. Это сходство, однако, лишь конвергентное, связанное с родом пищи.
У многочисленных мирмекофагов — ехидны (Monotremata), сумчатых мурашеедов (Myrmecoides fasciatus Marsupialia), муравьедов (Myrmecoiagidae, Xenarthra американские неполнозубые) и т. д. имеется длинный червеобразный язык, сходный и здесь только в силу конвергенции.
Как известно, у дятлов опорой подвижного и далеко высовывающегося языка являются необычайно длинные подъязычные кости. У одного из тритонов, способного выбрасывать свой длинный язык очень далеко, также имеется длинный подъязычный аппарат, весьма сходный с тем же элементом дятлов.
Итак, в условиях сходного образа жизни систематически далекие формы или их органы оказываются сходными.
Следует иметь в виду, что конвергентное сходство, обусловленное одинаковым образом жизни, все же поверхностно и не обманет современного морфолога и систематика.
Сходство их всегда строится на разной генетической основе. Например, столь сходные между собою сумчатый крот и златокрот все же сохраняют признаки различий, определяющие их принадлежность к различным систематическим группам. Этот вывод касается решительно всех случаев конвергенции.
Становится ясным, что конвергентное сходство между неродственными формами есть сходство, обусловленное существованием неродственных форм в одной и той же стации или сходством их образа жизни. Однако, если это так, то нам остается признать, что и в случаях дивергенции и в случаях конвергенции мы имеем дело по существу с одним и тем же процессом эволюции. В первом случае родственные формы стали несходными в различных условиях существования. Во втором — неродственные формы стали сходными в сходных условиях жизни. Но и для первого и для второго случая остается справедливым один общий вывод: наличие эволюционных преобразований, идущих по руслу приспособительной изменяемости.
Таковы, в кратких чертах, экологические доказательства эволюции.