Факты свидетельствующие об этом, весьма многочисленны, охватывая все данные биологии. Из их числа остановимся лишь на некоторых.
а. Прежде всего, данные показывают факт закономерной смены форм животного и растительного мира на протяжении последовательных геологических эпох. Всюду подчеркнуто угасание старых форм и возникновение новых. Таблица времён ничего не говорит о преемственной смене отдельных видов, зато в ней, при внимательном изучении, найдется материал, позволяющий заметить последовательное преобразование отдельных естественных групп. Так, в силуре происходит преобразование беззамковых плеченогих в замковые, Nautiloidea с прямыми раковинами заменяются формами с закрученными раковинами, цистоидеи уступают место бластоидеям, древние четырехлучевые кораллы — шестилучевым, на смену регулярным морским ежам появляются иррегулярные и т. п. Во многих подобных случаях палеонтологический материал позволяет явственно констатировать, что эта смена форм осуществляется в порядке изменения предшествующей организации и преобразования ее в новую. Другими словами, констатируется эволюция отдельных естественных групп организмов.
б. Нужно, однако, отметить, что изложенные факты еще не дают точных доказательств эволюции. Поэтому, хотя, например, Кювье своими личными исследованиями обнаружил факт смены форм и преобразования отдельных естественных групп в последовательных геологических горизонтах, он все же не делал отсюда эволюционных выводов и объяснял эту смену форм теорией катастроф. Для установления факта преемственной смены форм надо найти случаи, когда мы можем проследить последовательное преобразование серий родов и видов, в которых можно показать, что конкретные виды-потомки происходят через превращение видов-предков.
Классическим примером подобных случаев служит эволюция предков современной лошади, которую мы и рассмотрим.
Эволюция лошади охватывает большой промежуток времени. Древнейший предок лошади Eohippus относится к началу третичного периода, тогда как современная лошадь Equus известна только с плейстоцена, т. е. относится к четвертичному периоду.
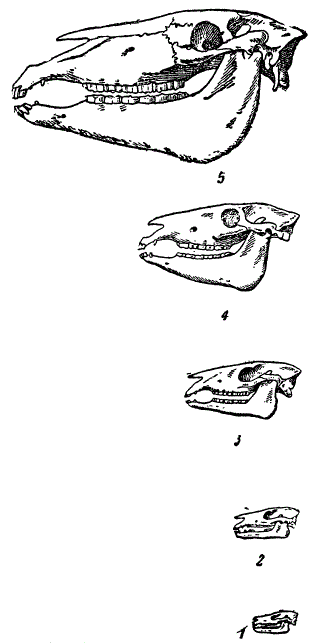
Черепа некоторых предков лошади, сопоставленные в одном масштабе. 1 — эогиппус, 2 — орогиппус, 3 — мезогиппус, 4 — меригиппус, 5 — лошадь (Из Мэтью)
Виды рода Eohippus были маленькими лесными животными, высотой в 30 см. Они имели по четыре пальца на передних и три на задних ногах, что облегчало хождение и бег по топкой почве лесных болот. Судя по неспециализованным зубам, питались эти животные мягкой растительной пищей. Относятся они к нижнему эоцену Северной Америки.
За этой формой следуют среднеэоценовые орогиппусы (Orohippus), у которого на передних ногах имеются еще развитые 4 пальца. В верхнем эоцене появляется эпигиппус (Epihippus), у которого 4-й палец явственно уменьшен.
В олигоцене жил потомок предыдущих форм — мезогиппус (Меsohippus). У видов этого рода — три пальца, но при этом средний палец (третий) заметно сильнее остальных. Рост достигает 45 см.
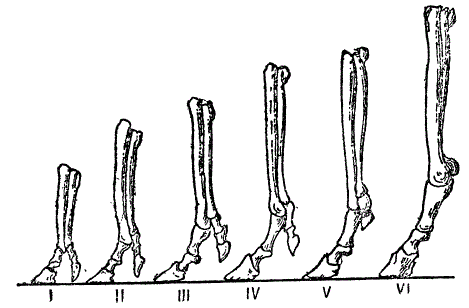
Профильное изображение передних конечностей ряда лошадиных предков. 1/5 естественной величины. 1 — Mlohippus Bairdii, средн. олигоцен, 2 — М. intermedius, верхн. олигоцен, 3 — Parahippus atavus, средний миоцен. 4 — Merichippus primus, средний миоцен, 5 — М. eohipparion, средн. миоцен, 6 — Pliohippus Lalianus, нижний плиоцен (Из Вебера)
В отмеченном ряду, в соответствии с изменением строения конечностей и роста, намечаются интересные изменения в зубной системе. Бугорчатые передние зубы эогиппусов, приспособленные, как сказано выше, к мягкой растительной пище, превращаются у эпигиппусов в зубы с бороздками (гребнями). Это достигается слиянием отдельных бугров. Эволюция захватывает и ложнокоренные зубы. У эогиппусов они имеют треугольную форму, у эпигиппусов они становятся похожими на коренные (моляры), приобретая четырехугольную форму. Этот процесс моляризации ложнокоренных зубов, зависимый от перехода к более грубой степной растительной пище и необычайно характерный для лошади, делает ложнокоренные зубы похожими на коренные (моляры).

Процесс моляразации ложнокоренных зубов у предков лошади. 1 — эогиппус, 2 — орогиппус, 3 — эпигиппус. У эогиппуса все ложнокоренные имеют еще треугольную форму (Из Вебера)
В верхнем олигоцене мезогиппус уступает место целому ряду форм: миогиппусу (Miohippus), анхитерию (Anchiterium), парагиппусу (нижний миоцен). Миогиппус и анхитерий жили в лесу, сохраняли сильно развитые боковые пальцы, служащие опорой на мягкой лесной почве. Напротив, парагиппус отличался более короткими боковыми (IV и V) пальцами и, повидимому, придерживался открытых пространств, обладая более быстрым бегом в более крупным ростом.
Первые две ветви вымерли бесследно, а парагиппус явился родоначальником следующего этапа лошадиного «ряда», — меригиппуса (Merichippus). Меригиппусы, несомненно, были жителями открытых пространств, причем у разных видов этого рода идет процесс укорочения боковых пальцев, у одних видов они длиннее, у других — короче, последние — прогрессивнее и ближе к быстроногим однопалым лошадям.
Наконец, у плиогиппуса (Pliohippus, плиоцен) этот процесс заканчивается. Плиогиппус — это древняя американская однопалая степная лошадь. Ее производят от меригиппуса. Последний, кроме того, является родоначальником гиппариона (нижний плиоцен), у которого имелось три пальца, из них второй и третий — рудиментарные. Гиппарион имел широкое распространение, проник даже в Евразию, но, повидимому, вымер бесследно.
В течение плиоцена плиогиппус дает еще одну однопалую форму — плезиппуса. По форме и размерам он уже близок к современной, известной с плейстоцена лошади Equus и, по данным американских исследователей (Мэтью) должен рассматриваться, как непосредственный предшественник ее. Возникнув в Америке, современная нам лошадь затем проникает в Евразию в числе нескольких видов. Что касается упомянутого американского плиогиппуса, то в верхнем плиоцене от него, кроме плезиппуса, отчленяется ряд других форм (гиппидиум, оногиппидиум и др.), присущих Южной Америке, которые затем вымирают. В конечном счете все американские лошади вымерли, а европейские уцелели и затем вторично попали в Америку, на этот раз завезенные сюда вместе с завоевателями-европейцами, в начале XVI в.
В общем, отношения между перечисленными выше формами могут быть представлены схемой, причем на ней показаны, кроме перечисленных форм и некоторые другие, образующие боковые, бесследно вымершие ветви. Таковы, кроме упомянутых анхитерия и миогиппуса, маленький гиракотерий, живший в эоцене и близкий к эогиппусу, с которым он связан, повидимому, общим происхождением.
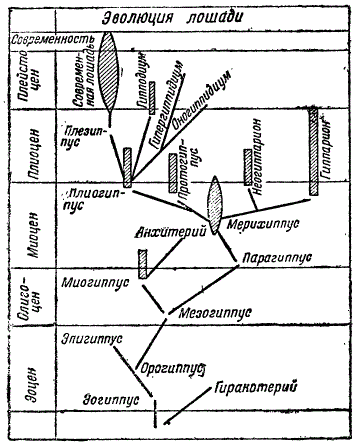
Схема эволюции лошади. По Мэтью, с сокращениями
Краткий обзор эволюции лошадей убедительно показывает процесс эволюции, ведущий к возникновению новых видов путем преобразования их предков.
Совершенно сходное заключение можно сделать при рассмотрении другого примера — эволюции слоновых (хоботных). В истоках эволюционного ряда слонов можно констатировать относительно мелкие формы, представленные жившими в олигоцене меритериями, достигавшими величины современных тапиров. Меритерии были еще далеки от слонов, в частности, по строению зубной системы. От них берут начало олигоценовые палеомастодонты, которые, в общем уже сильно напоминают слонов, в особенности по строению черепа. Рядом с палеомастодонтами следует поставить файомию (Phiomia), повидимому, боковую ветвь слонового ствола. От файомии происходит целый пучок форм — средне- и верхнемиоценовые трилофодоны, нижнеплиоценовые тетралофодоны и другие формы (вся правая часть), которые целиком вымерли.
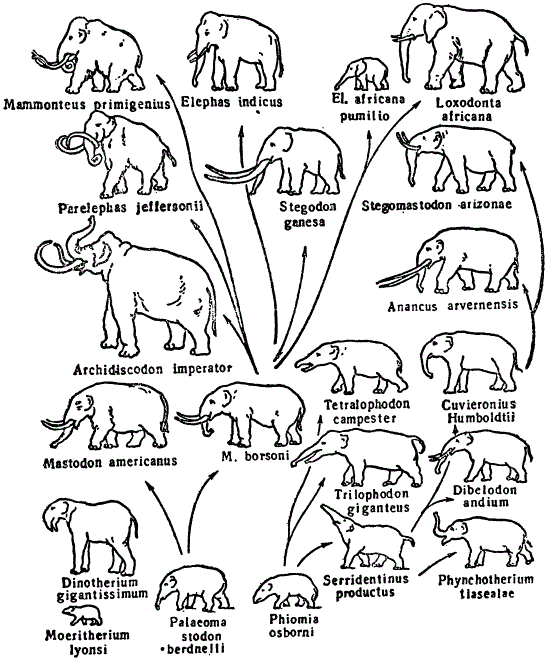
Эволюция хоботных (Из Берри)
Что касается палеомастодонтов, то их дальнейшая эволюция приводит к возникновению новых форм мастодонтов, которые рассматриваются, в свою очередь, как предки стегодонов. Последние образуют переход от мастодонтов к настоящим слонам, представленным в геологической современности вымершим мамонтом и индийским и африканским слонами.
Если проследить изменения, которые характеризуют современных слонов по сравнению с их предками, то можно констатировать следующие черты: 1) увеличился рост, 2) мощного развития достиг хобот, 3) развились резцы верхней челюсти, превратившиеся в бивни, 4) остальные зубы подверглись редукции, кроме коренных, которые сильно развиты у современных слонов; 5) число поперечных гребней на зубах у современных слонов увеличено. Так, у Palaeomastodon их три, у Mastodon этих гребней бывает пять и шесть, у стегодона имеется 6—12 низких, крышеобразно построенных гребней. Наконец, у современных слонов их до 27 (индийский слон). Изменились, конечно, и другие признаки. В целом, картина эволюции слонов явственно носит дивергентный характер, связана с вымиранием множества древних форм и вместе с тем приводит к усилению и накоплению новых изменений.
В обоих избранных примерах (лошади, слоны) явственно видно, что смена форм во времени носит преемственный характер и что лошадь и слоны являются современными нам этапами длительного процесса эволюции. Примеров этого рода мы знаем относительно большое число и некоторые филогенетические деревья отдельных естественных групп (например, головоногих моллюсков, сем. верблюдов, некоторых групп парнокопытных и т. д.) разработаны примерно с той же степенью полноты, которая достигнута в отношении сем. лошадиных и группы хоботных.
в. О факте преемственности в преобразовании форм свидетельствует также наличие так называемых переходных форм, под которыми понимаются формы, преемственно связывающие во времени две соседние группы — предшествующую и последующую.
Как правило, переходные формы и группы совмещают некоторые особенности соседних групп. Палеонтологи хорошо знали такие формы еще до Дарвина. Дарвин ссылается на Оуэна, называвшего их «обобщенными формами» и на Агассица, писавшего о «синтетических» типах. Примеры переходных форм довольно многочисленны. Остановимся лишь на некоторых из них.
1. Явственно промежуточное положение занимают, например, первоптицы (Archaeomithes) археоптерикс (Archaeopteryx) и археорнис (Archaeornis). В ряде признаков первоптицы сохраняют типичные черты пресмыкающихся. С этими последними первоптицы сближаются, прежде всего, по строению черепа. Так, у археорниса отсутствовал роговой чехол клюва, челюсти были вооружены зубами, перьев на коже головы не было. Скелет также в значительной степени был рептильным. Позвонки у первоптиц были подвижные, ребра очень тонкие, без характерных для птиц крючковидных отростков, брюхо было защищено, как у современных крокодилов, брюшными ребрами, которых у птиц не бывает. Хвост был очень длинным, как у ящериц. Скелет передних конечностей нес три свободных пальца, которые были вооружены длинными когтями, причем метакарпальные кости не были еще срощены, и, следовательно, кисть обладала свободной подвижностью.
Однако, с другой стороны, передние конечности и хвост первоптиц несли явственно развитое оперение — признак, свойственный только птицам. Несомненно, что археоптерикс и археорнис были плохими летунами. Некоторые авторы полагают, что они не летали вовсе, а только планировали. Другие считают, что первоптицы летали, но только на короткие расстояния (перепархивание с дерева на дерево). Это заключение основано на ряде данных. Так, крыло первоптиц по форме сходно с крылом современных порхающих птиц, в особенности золотого фазана. Его величина не значительна. Маховые перья у первоптиц не имели прочной опоры в скелете пястья (metacarpus). Длинный оперенный хвост также не вяжется с быстрым полетом. Первоптицы представляли только одну из первых стадий в историческом развитии полета. Описанные черты их организации отличаются известной двойственностью. Рептильная природа первоптиц сочетается с признаками птиц. Эти отношения очень хорошо подчеркивают переходный характер их морфологической организации — от пресмыкающихся к птицам.
2. Такое же положение промежуточного звена между пресмыкающимися и млекопитающими играют некоторые представители древней примитивной группы рептилий — звероящеров (Theromorpha). В облике этого пресмыкающегося много черт млекопитающих.
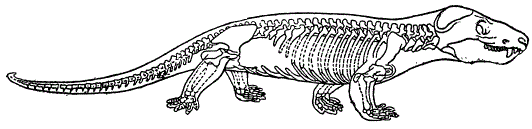
Циногнат (Из Ромера)
У пресмыкающихся конечности обращены в стороны от главной оси тела. Вследствие этого тело «висит» на них, как на боковых пружинах. У млекопитающих конечности прижаты к телу и подобраны под корпус; следовательно, последний опирается на них, а не подвешен к ним.
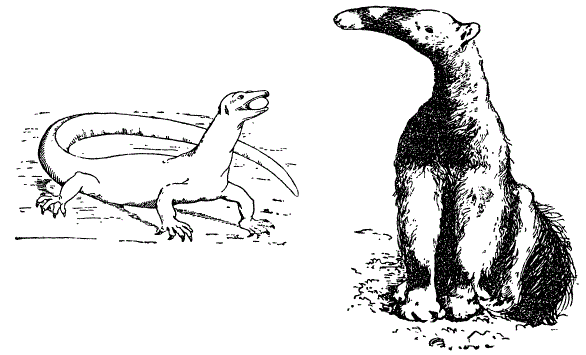
Положение конечности по отношению к туловищу у млекопитающих (молодой тамандуа-бандейра) и пресмыкающихся (серый варан). По разным авторам
В связи с этим локоть у млекопитающих обращен назад, а колено — вперед. Очень сходно положение конечностей у Cynognathus. Особенно интересно, однако, строение черепа у некоторых звероящеров. Число костных элементов в черепе некоторых из них уменьшается, приближаясь к черепу млекопитающих. Процесс этот происходит за счет редукции или слияния некоторых черепных костей. Так, например, у звероящеров происходит срастание заднетеменных костей, причем за их счет образуется характерная для млекопитающих межтеменная кость. Скуловая дуга образована за счет тех же элементов, что у млекопитающих. Далее, у циногнатуса явственно развито твёрдое нёбо — признак млекопитающих. Наконец, важным элементом их характеристики является развитие двух затылочных сочленовных бугров (как у млекопитающих) вместо одного, свойственного рептилиям. Зубной аппарат звероящеров диференцирован наподобие млекопитающих (наличие диференцированных резцов, клыков и коренных зубов).
Наряду с этим у звероящеров сохраняются признаки рептилий, например, теменное отверстие в черепе, ряд черепных костей, свойственных рептилиям, и т. п. Таким образом, звероящеры, могут рассматриваться как переходные формы, связывающие рептилий с млекопитающими.
Количество переходных форм велико, и они подтверждают наличие связей между соседними естественными систематическими группами животных.
3. Среди растений также можно указать на ряд форм, обладающих чертами переходной организации. Таковы, например, семенные папоротники — птеридоспермы, связывающие папоротникообразные с семенными. Эти растения, игравшие значительную роль в каменноугольном периоде, производили не только споры (как папоротники), но и семена, как семенные растения. Хотя вайи птеридосперм были похожи на вайи папоротников, с другой стороны, они напоминали рассеченные листья некоторых семенных. Проводящая ткань их стеблей анатомически сходна с древесиной голосеменных (саговников) и т. п.
Перечисленные выше факты устанавливают явление филогенетической преемственности между видами (эволюция лошадиных, эволюция хоботных) и между соседними естественными группами (пресмыкающиеся — птицы, пресмыкающиеся — млекопитающие). Подобных примеров, почерпнутых из области палеонтологии, известно относительно много, и в их свете наличие филогенетической преемственности между последовательно развивающимися формами и соседними группами — твердо доказанный факт.
г. До сих пор мы имели дело с палеонтологическими данными, позволяющими видеть эволюционный процесс, как реальную форму исторического развития. Палеонтологический материал, однако, освещает процессы эволюции не полно. Еще Дарвин отмечал неполноту палеонтологических документов. Они — фрагментарны, отрывочны. Формы, не имеющие скелетов или раковин, в большинстве случаев не сохраняются совсем. Палеонтолог сплошь и рядом имеет дело с неполными скелетами или даже с отдельными, разрозненными костными элементами. Организм, как целое, вообще недоступен ему. Кроме того, палеогеология доказывает лишь прошлое эволюционного процесса. Мы же должны научиться видеть его и в геологической современности, установить, что эволюция не только происходила, но и происходит ныне. Только при этом условии становятся вполне убедительными и палеонтологические данные.
Многочисленные факты позволяют вскрыть явление эволюции не только при изучении палеонтологических данных, но и иными путями.
г. Эволюционные выводы вытекают, прежде всего, из сравнительно-анатомического изучения современных нам форм. Выше мы уже касались явления специальной гомологии, чтобы подчеркнуть черты сходства между формами одной естественной группы и соответственно — идею монофилии. Однако, с эволюционной точки зрения, важно отметить не только черты сходства в строении гомологичных органов, но и различия между ними. В самом деле, если можно показать, что гомологичные органы становятся различными, то самый факт возникновения различий между ними приобретает значение доказательства эволюции.
Чтобы в этом убедиться, проанализируем ранее использованный пример специальной общей гомологии — передние правые конечности ряда позвоночных. Легко констатировать, что все эти конечности, сходные в общей архитектонике (единство типа организации), в то же время весьма различны в деталях. Так, передняя конечность кита образуется за счет того же числа гомологичных элементов, что и крыло птицы, но отличается от последнего в ряде частных признаков. Это связано с тем, что кит приспособлен к плаванию, а птица — к полету. Правые передние конечности этих животных гомологичны друг другу, но в то же время изменены, т. е. очевидно эволюировали в разных направлениях. Стало быть эволюировали и их обладатели.
Еще явственнее это в случаях неполной специальной гомологии. Нога лошади характеризуется выпадением ряда костных элементов. Так, в ней исчезла лучевая кость (от нее сохранился лишь рудиментарный элемент — olecranon), а в пясти и запястьи — костные элементы I, II, IV и V пальцев. Сильно развит только третий палец, да остались костные элементы, связанные с исчезнувшими вторым и четвертым пальцами (две ossa calamiformia). Легко показать, что эти отношения — прямой результат эволюции. Так, у эмбрионов лошади имеются и лучевая кость и полные костные элементы трех пальцев (II, III и IV). Иногда в качестве аномалии, сохраняется и лучевая кость. Эти факты свидетельствуют о том, что лошадь произошла по крайней мере от трехпалого предка. Сопоставляя этот вывод с данными палеонтологии, мы с полным основанием приходим к выводу, что лошадь — продукт эволюции многопалых предков.
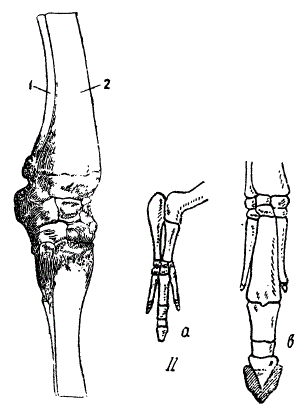
Гомологизация передней конечности лошади с типичной конечностью. I — сравнительно-анатомические данные. Случай развития локтевой кости у годовалого жеребенка. 1 — локтевая. 2 — лучевая кость. II — эмбриологические данные: а — нога шестинедельного зародыша лошади. Показана редукция четвертого и пятого пальцев, в — дальнейший ход процесса (Из Вебера и Полякова)
Не менее убедительны доказательства эволюции, основанные на анализе случаев общей гомологии. Общая гомология охватывает органы, повторяющиеся в теле одного и того же организма, как вдоль длинной оси его тела, так и вдоль поперечных осей его. Дарвиниста интересуют по преимуществу различия, возникающие в случаях общей гомологии. В виду этого, рассмотрим некоторые примеры общей гомологии.
1. Гомотипия — развитие двух органов сходного плана, противостоящих друг другу, как зеркальное отражение. Таковы, например, правая и левая рука, правый и левый глаз и т. п. Гомотипичнце органы симметричны друг другу и притом энантиоморфны, т. е. относятся друг к другу как зеркальное (обращенное) отражение. Однако симметрия их относительна и часто переходит в асимметрию, которая у многих форм становится их видовым признаком. Так, например, у краба Uca pugilator, живущего в мангровых зарослях южной Кубы, правая клешня значительно меньше левой. При приближении наблюдателя краб залезает в норку и прикрывается большой клешней. Так как для гомотипических органов характерна симметрия (как правиле, правая и левая клешни одинаковы), ясно, что виды, обладающие асимметричными гомотипическими органами, произошли от видов с симметричным расположением их. Следовательно, явление асимметрии гомотипических органов имеет вторичное происхождение и является результатом эволюции. Как видно из приведенного примера (краб), асимметрия, являясь видовым признаком, имеет в то же время приспособительное (в нашем примере — защитное) значение.
2. Гомодинамия (сериальная гомология). Гомологичные органы, расположенные по длинной оси тела в сериальном порядке и развивающиеся из сходных зачатков, называются гомодинамичными.
Примеры: позвонки позвоночного столба, серия ребер, аортальные дуги, серия ножек у ракообразных и т. д.
Примером могут служить конечности речного рака. Отдельные конечности его следуют друг за другом и сходны в общем плане, состоя из равного числа частей, но резко различны в деталях строения. Первая и вторая пара конечностей функционируют как антенны, ряд пар конечностей преобразован в ротовые органы, далее следуют так называемые ногочелюсти, при помощи которых рак держит свою добычу, пара хватательных клешней, пять пар ходильных ног, столько же брюшных ножек, обеспечивающих (быстрыми колебаниями) приток свежей воды к жаберным карманам и придерживающих у самок отложенную икру, наконец, одна пара широких хвостовых плавательных конечностей. Достаточно сравнить эту последнюю пару с парой клешней, чтобы убедиться, какие глубокие изменения претерпевают гомодинамные органы, в связи с различием их функций.
3. Гомономия — явление сходства частей органа, расположенных по его поперечной оси. Пример: пальцы рук — гомономны друг другу.
И в этом случае наблюдаются резкие приспособительные изменения, имеющие значение видовых особенностей. Второй палец задней ноги у лемура Perodicticus calaberensis, например, резко отличается от остальных.
Вместо обычных трех фаланг, он содержит костный пенёк. Вооруженный острым когтем (в отличие от ногтей на остальных пальцах), он выполняет функцию «пальца для очистки меха». Совершенно ясно, что подобное строение пальца — результат эволюции.
Сказанное выше прекрасно иллюстрируется и на растительном материале. Рисунок показывает, какие глубокие превращения претерпевает лист, принимающий на себя самые разнообразные функции. Возможность наблюдать направления и характер эволюции гомологичных органов придает их изучению большую ценность. Превращения гомологичных органов отображают не только ход эволюции этих последних, но и эволюцию их обладателей (биологических видов).
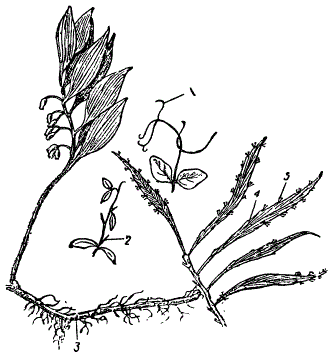
Гомология у растений. 1 — усики посевного гороха — гомологи листьев: 2 — иглы у барбариса — гомологи листьев. 3 — корневище купены, гомологичное стеблю: 1 — часть стебля филлантуса. Кладодии (4) гомологичны стеблю, а не листьям
д. Напомним, наконец, явления аналогичных сходств, под которыми понимаются случаи возникновения сходства между органами, генетически не имеющими ничего общего. Таково, например, сходство между спинным плавником акулы и кита, крыла насекомого и крыла птицы, глаза какого-либо кольчатого червя и млекопитающего и т. п. Возникновение подобных аналогичных сходств может быть понято только с исторической точки зрения, т. е. как результат соответственных эволюционных преобразований различных органов.
е. Остановимся теперь на явлении рудиментации органов. Под рудиментами понимаются органы, которые нормально закладываются, но в дальнейшем перестают развиваться и остаются у взрослых форм в недоразвитом состоянии. Такой рудиментарный орган утрачивает свою нормальную функцию, приобретает новую или не несет какой-либо специальной функции. Такие органы и называются рудиментами. Приведем некоторые примеры рудиментарных органов.
1. Рудиментация пальцев. Распространеннейшей формой ее следует считать рудиментацию пальцев у наземных позвоночных, в особенности у птиц и млекопитающих. У многих птиц, например, гагар (Colymbidae), поганок (Podicipidae) и др. рудиментирован II палец, заметно уменьшенный по сравнению с остальными. В крыле птиц пальцы частично полностью исчезли, частично рудиментированы. У млекопитающих рудиментация пальцев широко распространена, характеризуя большие систематические группы.
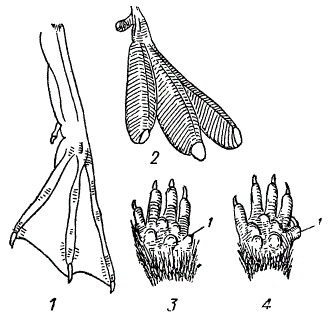
Рудиментарные органы. 1 — лапка гагары; 2 — лапка поганки с рудиментарный первым пальцем (1); 3 — лапка полевки (Microtus arvalis); 4 — она же случай ненормально развитого первого пальца (1). (Из Житкова)
У полевок (Microtus) полностью рудиментирован I палец передних ног; у собак имеется рудиментарный и не функционирующий I палец; у свиней захвачены процессом рудиментации II и V пальцы, тогда как III и IV служат опорой ноги; у лошадей полностью рудиментированы все пальцы, кроме III; у вымерших предков лошади, начиная с парагиппуса, нарастает рудиментация IV и II пальцев; интересные формы рудиментации пальцев наблюдаются у лемуров.
2. Рудиментация конечностей — также частое, явление. У некоторых хвостатых амфибий рудиментированы и передние и задние ноги, причем степень рудиментации ног прямо пропорциональна длине тела. Очень удлиненные формы имеют резко рудиментированные конечности. Те же отношения у ящериц; например, у желтопузика, имеющего змеевидную форму тела, имеются маленькие рудименты задних ног. У питонов явственно выражены рудименты задних конечностей.
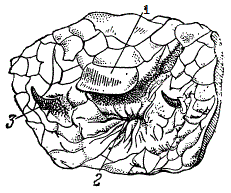
Рудименты конечностей у питона (Python reticulatus). 1 — анальный щиток. 2 — анус, 3 — рудименты конечностей (По препарату Биол. Музея Акад. Комм. Поспит., из Общей биологии Дорфмана)
Среди млекопитающих у китов рудиментированы задние конечности и тазовый пояс, причем у одного и того же вида степень их рудиментации колеблется. У дельфинов (зубастые киты) задние конечности полностью исчезли. Однако, и здесь можно доказать, что их исчезновение — лишь конечный результат рудиментации. Так, было показано, что у эмбрионов дельфинов закладки конечностей совершенно явственны, но затем они исчезают. В одном случае у взрослого Delphinus delfis были обнаружены явственные рудименты задних конечностей.
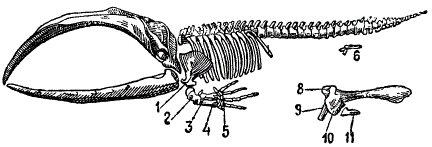
Рудименты тазового пояса и конечностей у кита (Baluena mysticetus). 1 — лопатка, 2 — плечо, 3 — лучевая к., 4 — локтевая к., 5 — кисть, 6 — рудимент таза, представленного ниже, 7 — подвздошная к., 8 — лобковая, 9 — седалищная, 10 — бедряная, 11 — большая берцовая. (Из Вебера и Абеля)
3. Рудиментация зубов — также частое явление. У муравьедов они полностью рудиментированы, у остальных неполнозубых (броненосцы) зубы лишены эмали и корней. У хищных Carnivora заметно рудиментированы резцы, а у части парнокопытных — клыки и т. п.
4. Рудименты у беспозвоночных также многочисленны. У двукрылых рудиментирована задняя пара крыльев, превращенных в так называемые галтеры или жужжала. Нет задних крыльев у некоторых клопов, например, у красного клопа. У некоторых жуков (Carabus, Calosoma и др.) задние крылья заметно рудиментированы. У стафилинид сильно рудиментированы элитры. У самок светляка крыльев нет вовсе. Более или менее полная рудиментация крыльев типична для многих островных насекомых из различных отрядов (Diptera, Coleoptera, Lepidoptera и т. д.). У многих форм, в том числе у пещерных, рудиментированы глаза. Наблюдается также рудиментация ротовых органов (например, у поденок, у некоторых бабочек и т. д.). В различных группах червей, например, у мермитид (Nematodes) и т. д., наблюдается рудиментация кишечника.
5. Рудиментация у растений касается всех органов. У некоторых грибов (Saprolegnia, Achlya) наблюдается рудиментация антеридиев. В цветах Меlandrium рудиментированы тычинки, у многих зонтичных недоразвита чашечка, у самоопылителей наблюдается рудиментация венчика. У некоторых растений стебель берет на себя функцию листьев, а листья рудиментарны и т. д.
Рудименты представляют двойной интерес. Во-первых, рассмотренные выше признаки не являются уродствами (в пределах данной группы организмов) и имеют значение важных систематических признаков соответствующей естественной группы. Во-вторых, сличение рудиментов с их нормальными гомологами (например, рудиментированного тазового пояса с нормальным) показывает, что рудиментарный орган есть орган изменившийся, эволюировавший. Это последнее положение наиболее для нас важно. Особенно наглядны случаи, когда у массы представителей данного вида какой-либо орган рудиментирован, но и у отдельных особей того же вида он развивается полностью.
На рисунке показана передняя лапка полевки Microtus arvalis, нормально имеющей четыре пальца, а именно II, III, IV, тогда как I палец явственно рудиментирован. Б. М. Житков описал случай, когда у полевки того же вида на месте рудимента оказался развитым нормальный I палец. Этот случай превосходно показывает, что рудиментация I пальца у полевки — результат изменения, ставшего видовым (даже родовым) признаком.
Подобные случаи не редки. К ним относятся, например, развитие у лошади трех пальцев — нормального III и двух боковых (II и IV), соответствующих грифельным костям нормальной лошади и т. д.
Описываемые случаи, известные под названием атавизмов, рассматриваются как возврат к признакам предков, или анцестральным признакам.
Случаи появления развитого I пальца у полевки, боковых пальцев у лошади или задних крыльев у красного клопа доказывают, что рудименты — результат изменения соответственных нормальных органов, а стало быть, виды с рудиментированными органами произошли от форм, обладавших нормальными органами. Явление рудиментации — одно из доказательств эволюции.
ж. Ценные доказательства эволюции дает анализ многочисленных биогеографических отношений между различными видами и их группами.
Из большого числа биогеографических фактов, доказывающих эволюцию, остановимся лишь на некоторых. Прежде всего остановимся на сравнительной характеристике некоторых зоогеографических областей. Сравним сперва фауны Эфиопской и Индо-Малайской областей. Как в первой, так и во второй обитают человекообразные обезьяны, обеим областям свойственны полуобезьяны лори (семейство Lorisidae, подсем. Lorisinae), ящеры (отряд Pholidota), виверры (сем. Viverridae), кошки (сем. Felidae), гиены (сем. Hyaenidae), слоны (отр. Proboscidea), свиньи (сем. Suidae), оленьки (подотр. Tragulidea), антилопы (Antilopinae), буйволы, носороги и т. д.
Чем объясняются черты зоогеографической общности этих областей? Очевидно, вопрос идет об исторических связях между Африкой и Южной Азией и, следовательно, о кровном родстве общих им элементов фауны.
Фактическое положение вопроса таково. В третичное время фауны Африки и южной Азии обладали замечательным сходством. Так, в области Сиваликских холмов (подножье Гималаев) в плиоценовых слоях были обнаружены костные остатки ряда животных, частично ныне обитающих в Африке, частично в южной Азии. Таковы макаки (южная Азия), палеопитек (вымер), дриопитеки (вымерли), тонкотелые обезьяны (южная Азия), мастодонты, стегодоны (вымерли), носороги, бегемоты (Африка), свиньи (африканского типа), жирафы, антилопы (Африка) и т. д. Сходная фауна найдена в плиоцене Европы и Египта. Так, дриопитеки были широко распространены не только в южной Азии, но и в Европе. Этот род составляет, повидимому, корень, от которого берут начало современные африканские человекообразные обезьяны (горилла и шимпанзе) и предки человека.
Из этих данных видно, что эфиопская и индо-малайская фауны имеют несомненный общий корень. Отсюда сходство их фаунистических элементов. Однако отмеченные черты сходства следует рассматривать в их связях с различиями. Обеим областям свойственны общие отряды и иногда семейства, но роды и виды, а часто и семейства различны.
Так, например, человекообразные обезьяны Африки (гориллы и шимпанзе) образуют особые роды, в значительной степени отличные от индо-малайского оранга. Индо-малайские носороги Dicerorhinus и Rhinoceros принадлежат к другим родам, чем африканские Diceros и Ceratotherium, и хорошо отличаются от последних по ряду признаков. Африканские речные свиньи Potamochoerus и бородавочник Phaecochoerus принадлежат к другим родам, чем индийские Sus. То же следует сказать об антилопах и оленьках, об африканском (Loxodonta africana) и индийском (Elephas indicus) слонах, об ящерах и т. д.
Подобные отношения весьма обычны. Почему же именно, как правило, в разных зоогеографических областях одни и те же группы форм оказываются представленными разными видами, родами и даже семействами? Научное исследование не может, разумеется, ссылаться на божью волю.
Историческая точка зрения дает вполне удовлетворяющий ответ на эти вопросы. В геологическом прошлом, как указывалось, имел место тесный контакт между элементами африканской и южноазиатской фаун. Распространившись и там и здесь, соответственные формы в дальнейшем эволюировали независимо одна от другой. Геологическая давность процесса пространственного разобщения африканских и южноазиатских представителей одной и той же формы и привела к тому, что они обособились не только в различные виды, но и в разные роды.
Таким образом, например, африканский и индийский слоны должны рассматриваться как разобщившиеся кровно-родственные формы, берущие начало от далекого общего корня, а затем эволюировавшие, с одной стороны, в Африке, а с другой, в Индии независимо друг от друга и достигшие родовых различий. Действительно, семейство слонов (Elephantidae) берет начало от стегодонов (Stegodon), широко распространенных еще в плейстоцене южной и восточной Азии. От стегодонов возникают две линии. Одна — индийская — ведет к современному индийскому слону, другая — к африканскому, который и ныне сохраняет некоторые черты сходства со стегодонами. Из этих фактов видно, что обособление африканской линии слонов от южно-азиатской — очень давнее, почему их признаки и достигли степени родовых различий.
Обратимся теперь к сравнению фаун Евразии и Северной Америки. Здесь мы найдем существенно иную картину, а именно — громадное число близко сходных видов. Таковы, например, среди млекопитающих: американский и европейский бобры, далее пеструшки, зайцы, норки, выдры, лисицы, песцы, медведи, россомахи, рыси, северные олени, лоси. Многие из перечисленных форм безусловно принадлежат к одним и тем же видам (россомахи, рыси, бобры, северный олень, лось и др.), остальные — близкие виды.
Почему на этих двух материках много близких или даже идентичных видов? Исторический взгляд и в этом случае дает ответ. Очевидно, фауны Евразии и Северной Америки обособились геологически недавно — такое объяснение легко подтверждается фактами. Разобщение Азии и Северной Америки — очень позднее геологическое явление, относящееся к концу ледникового периода. Следовательно, близкие виды обоих материков разобщились геологически настолько недавно, что процесс их взаимного расхождения не достиг большой глубины.
Значит, чем позднее произошло разобщение материков, тем в меньшей степени успели разойтись родственные формы, тем больше сходство между ними. Наоборот, чем раньше произошло это разобщение, тем глубже степени расхождения форм соответственных материковых фаун, тем меньше сходство между ними. Итак, сопоставление фаун зоогеографических областей убеждает в том, что под влиянием указанных выше причин произошло ослабление или даже утрата сходства, т. е. процесс эволюционных преобразований, приведший к возникновению новых видов, все же связанных узами родства с соответствующими видами других зоогеографических областей.
з. Подобные же выводы могут быть сделаны в результате анализа фауны океанических островов, удаленных от материковых массивов. Островная фауна и флора характеризуются, как правило, большим числом эндемичных видов.
Под эндемиками понимаются формы, свойственные ограниченным местообитаниям или географическим пунктам (бассейн определенной реки, озера, горный массив, остров и т. п.), за пределами которых данные формы не встречаются. Эндемизм широко распространен в самых различных группах растений и животных. Эндемики особенно интересны, когда возможно их сравнение с соответствующими, близкими им формами соседних областей, поскольку такое сравнение позволяет обнаружить, что эндемики — это измененные потомки и родственники других, неэндемичных форм. В этих случаях эволюция Еесьма отчетлива.
На Дарвина сильное впечатление произвели эндемики Галапагосских островов. Здесь особенно ярко проявился тот факт, что эндемичные формы — результат эволюции. Отношения, существующие на Галапагосских островах, необычайно интересны и в частности некоторые особенности их орнитофауны.
Галапагосский архипелаг расположен в Тихом океане, на экваторе, в 700 км от берегов Южной Америки. На островах зарегистрировано 76 видов птиц. Среди них 12 видов — хорошие летуны, распространенные очень широко, свободно пересекающие морские пространства между островами и материком, и они не отличаются от своих материковых собратьев. Таков, например, рисовый скворец.
Среди прочих галапагосских птиц имеется целый ряд эндемичных видов и даже эндемичных родов, которые живут только на Галапагосских островах и неспособны сообщаться с материком. Таковы: галапагосский канюк, галапагосская сова, галапагосские пищухи, галапагосские вьюрки Geospiza и др. В общем 85% галапагосских птиц — эндемики. Все эти формы изменены по сравнению с материковыми, образуя особые виды и роды.
Эти факты необычайно четко указывают на наличие эволюционного процесса. Если птицы свободно сообщаются с материком, а следовательно, и скрещиваются с материковыми формами, они сохраняют «материковую» видовую принадлежность. Если они осели на островах и утеряли связь с материком, они оказываются измененными. Сохраняя печать американского происхождения, они обособляются при этом либо в особые виды американских родов, либо даже — в особые роды, все же сохраняющие известные черты сходства с американскими видами. Факт эволюции видов, их пространственного превращения, здесь налицо.
Другая группа фактов касается процессов изменяемости форм уже в пределах галапагосской орнитофауны. Дарвин заметил, например, что галапагосские виды вьюрков Geospiza диференцированы по отдельным островам этого архипелага, различаясь преимущественно по форме клюва — начиная от тонкоклювых и кончая формами с относительно толстым клювом. В общем, на Галапагосских островах известно 32 формы вьюрков, по несколько видов на одном и том же острове.
Дарвин был поражен этой изменяемостью галапагосских вьюрков. В «Дневнике изысканий» он делает единственно возможный вывод из этих фактов, высказывая допущение, что все эти местные островные формы вьюрков — результат изменяемости (модификации) одного первоначального вида. Действительно, это допущение остается единственным объяснением описанных фактов.
Касаясь фауны Галапагосских островов, Дарвин писал: «В песне пересмешника, в хриплом крике стервятника, в больших, похожих на канделябры, опунциях (кактус. А. П.), я ясно видел соседство Америки, хотя острова эти и разделялись таким огромным количеством миль океана… Еще удивительнее было то обстоятельство, что большинство обитателей каждого отдельного островка этого небольшого архипелага отличалось между собою видовыми признаками, хотя и обнаруживало чрезвычайно близкое сродство (например, виды вьюрков. А. П.)… Я часто задавал себе вопрос, каким образом могли появиться эти столь оригинальные животные и растения; самый простой ответ на это, казалось, был тот, что обитатели различных островов произошли друг от друга и подверглись постепенным изменениям в ряду последующих поколений, и что все население архипелага произошло от особей большого материка, именно Америки». Дарвин указывал, что это допущение несравненно обоснованнее, чем предположение, что на каждом из этих пустынных и скалистых островков имел место специальный акт творения.
Этот взгляд нашел полное подтверждение. Как правило, эндемичная фауна и флора океанических островов несет отпечаток фауны и флоры смежного с ней материка. Этот факт явственно указывает на источник островной фауны и флоры. С другой стороны, эволюция островных форм доказывается тем, что, как правило, чем дальше океанические острова от берегов, тем больше на них эндемиков.
Так, например, фауна и флора Британских островов, геологически недавно обособившихся от Европы и, кроме того, близко к ней расположенных, очень сходна с европейской. Огромное большинство видов растений, наземных моллюсков, насекомых, амфибий, рептилий, птиц и млекопитающих принадлежит к европейским видам и едва отличимо от них.
Иная картина на более отдаленных от материков островах. Так, на Мадагаскаре отсутствуют обезьяны, антилопы, зебры, жирафы, слоны, носороги, львы, гиены и др. Геология острова не дает никаких указаний на их прошлое пребывание здесь. Мадагаскар отделился от Африки в верхнем миоцене, еще до того, как указанные выше формы могли проникнуть на него. Зато древняя фауна Мадагаскара, благодаря островной изоляции, сохранила здесь свои примитивные черты. Таковы мадагаскарские тенреки Centetinae, принадлежащие к наиболее примитивным насекомоядным, примитивные хищники из семейства виверр, в особенности фосса Cryptoprocta ferox, многочисленные лемуры и т. д. Давность обособления этих форм привела к тому, что они резко обособились от африканских и систематически. Так фосса выделяется в особое подсемейство Cryptoproctinae, в которое входит лишь один вид. Точно так же мадагаскарский лемур ай-ай так резко отличается от других лемуров, что его выделяют в отдельное подсемейство Chiromyinae. Впрочем, и другие мадагаскарские лемуры — Lemurinae и Indrisinae — весьма своеобразны и образуют особые подсемейства.
Таким образом, в случаях, когда острова изолированы от материка, их фауна сохраняет печать общности, но в то же время значительно отличается систематически.
В виду этого на океанических островах число эндемиков прямо пропорционально их удаленности от материка. На островах, изолированных от материков, образовались свои формы, преемственно связанные с материковыми. Эволюция здесь несомненна.
е. Однако, чтобы убедиться в этом, нет надобности непременно искать следов эволюции на островах. Напротив, не менее или даже более глубокие свидетельства ее видны и на широких пространствах материков. Любой вид, занимающий широкий ареал распространения, как правило, диференцируется на ряд связанных переходами местных рас или подвидов. Это явление — правило, отсутствие местных рас — исключение. Например, среди наших млекопитающих лишь немногие не дают подвидов, и притом все это — формы, имеющие небольшой ареал, отличающийся однообразием условий.
Подавляющее число форм млекопитающих, птиц и других групп животных, а также растений дает ряд географических рас. В пределах СССР примером может служить распространение лисицы Vulpes vulpes, которая встречается по всему Союзу, обитает, следовательно, в разнообразных условиях и соответственно им дает свыше 20 подвидов. При этом, если соседние подвиды отличимы в немногих признаках, то крайние подвиды резко отличны. Например, северная лиса Vulpes vulpes vulpes. обитающая на крайнем севере Европейской части Союза, имеет относительно крупные размеры, короткие уши и яркий густой мех, тогда как казахстанская лиса V. v. karagan, караганка, отличается малым ростом, значительно меньшей массой тела, длинными ушами и заметно более бледным и менее густым мехом. При этом, если разложить на столе серию шкурок подвидов лисиц — от северной V. v. vulpes к среднерусской V. v. crucigera, от нее к степной V. v. stepensis и далее вплоть до караганки, — получится серия, движение которой покажет развитие вида в пространстве.
Очень наглядно изменение вида в пространстве на примере географических рас барсуков Meles meles.
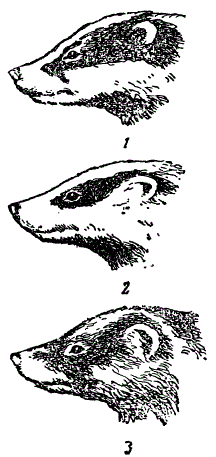
Подвиды барсуков. 1 — европейская форма, 2 — песчаный барсук, 3 — амурский (Из Бобринского)
Барсук имеет широкий ареал распространения (русская равнина, Кавказ, Крым, большие пространства Евразии — до Амурского края включительно). На пространстве этого обширного ареала он диференцировался на целый ряд форм, различающихся по многим признакам, в частности, по окраске меха, ширине темной полосы, идущей от глаза к уху, величине черепа.
Необычайное многообразие географических форм, отличающихся друг от друга главным образом по окраске и рисунку оперения, дает обыкновенный фазан Phasianus colchicus.
Рисунок показывает изменение формы клюва у географических рас Камышевой овсянки Emberiza schoenicla, причем наряду с изменением клюва изменяются размеры и масса тела. «Если поставить рядом маленькую камышевую овсянку с побережья Белого моря и огромную толстоклювую — с Сыр-Дарьи, то вряд ли придет кому-нибудь в голову считать их за один и тот же вид» (Портенко, 1928). И, действительно, крайние формы клюва различаются столь резко, что счесть их за клювы одного и того же вида можно только при наличии промежуточных звеньев (2—7).

Изменение формы клюва у подвидов камышевой овсянки (По Портенко)
Подобный же процесс пространственной местной диференцировки вида наблюдается и в других группах животных, например, у насекомых, у моллюсков и т. д., а также у растений.
Наиболее замечательной чертой пространственной диференцировки вида на местные географические формы является тот факт, что в результате ее ареал вида превращается в мозаику подвидовых ареалов.
Подвидовые ареалы в пределах общего ареала одного и того же вида, как правило, не совпадают, и, следовательно, подвиды (одного и того же вида) взаимно исключают друг друга в своих подвидовых ареалах. Такие взаимно исключающие друг друга географические расы, или подвиды одного вида называются викарными географическими расами.
Из сказанного ясно, что каждая викарная форма есть очевидный продукт ее ареала. Пересекая ареал вида, проникая из одного подвидового ареала в другой, просматривая серии викарных форм, можно убедиться, что вид изменяется, диференцируясь на подвиды, которые в дальнейшем обособляются в молодые новые виды. В ряде случаев мы можем наблюдать различные фазы этого процесса, которые мы, однако, рассмотрим позднее, в гл. X. Пока же лишь отметим, что подобно подвидам, молодые виды должны быть связаны между собою узами родства, — общей преемственной связью с видом-родоначальником. И действительно, например в отношении двух видов хорьков — темного и светлого (Putorius putorius и Р. ewersmanni) не трудно установить наличие близкого родства между ними, вследствие чего не всегда легко провести точную границу между подобной парой видов. Оба хорька дают нам пример начальной фазы взаимного обособления видов. В данном примере мы являемся очевидцами и современниками процесса обособления двух новых видов, возникающих от одной первоначальной формы и связанных между собою родством, как два брата, возникших от общего предка. Таких примеров известно большое число.
Итак, сравнительно-анатомические, палеонтологические и биогеографические данные, в их совокупности, позволяют нам значительно точнее установить самый факт эволюционного процесса. Мы констатируем его и в последовательных геологических горизонтах и в геологической современности, т. е. и в прошлом и в настоящем.