Как известно, дыхательная система растений построена по принципу множественности, т. е. на любом этапе дыхания принимают участие два фермента и более, выполняющих одну и ту же функцию.
Значение этого явления становится понятным, если принять во внимание температурный оптимум действия ферментов и сродство их к кислороду. Так, цитохромоксидаза, ферменты флавиновой группы и Cu-протеиды проявляют неодинаковую активность при различном содержании кислорода в тканях. Цитохромоксидаза обладает наибольшим сродством к кислороду, она активна при содержании его в среде 1—2%. Активность же ферментов флавиновой группы возрастает с увеличением концентрации кислорода.
В этой связи интересно, что наружные, хорошо аэрируемые ткани плода мандарина имеют кислородный оптимум при 21%, а внутренние, плохо аэрируемые, — при 10%. Ткани же мякоти, доступ кислорода к которым сильно затруднен, наиболее интенсивно дышат при содержании 5% кислорода в среде. Соответственно этому в плоде распределяются ферменты: в наружных тканях преобладает флавиновая группа (аэробные дегидрогеназы), в средних — медьсодержащие протеиды (дифенолоксидаза, аскорбиноксвдаза), во внутренних — цитохромоксидаза. Указанные ферменты отличаются и по отношению к температуре. Цитохромоксидаза функционирует при более высокой температуре, чем флавинопротеиды, имеющие низкий температурный коэффициент активности (Q10). Благодаря смене ферментативных систем создается возможность для осуществления дыхания в изменившихся условиях.
В ходе онтогенеза растений ферментативные процессы не остаются постоянными. Показано, например, что при созревании плодов яблонь Fe-протеиды сначала заменяются на Cu-протеиды, а затем на более «холодостойкие» флавиновые ферменты. Интересны наблюдения на хвойных. Известно, что хвоя лиственницы на зиму опадает, а хвоя сосны — нет. В формирующейся хвое лиственницы преобладает дифенолоксидаза, в период ее активного роста — цитохромоксидаза и пероксидаза. К осени активность пероксидазы в хвое лиственницы, постепенно падает. В хвое же сосны к осени активность пероксиаазы и дифенолоксидазы непрерывно возрастает, а зимой повышается активность ряда дегидрогеназ (у лиственниц они инактивируются). Полагают, что высокая активность дегидрогеназ способствует повышению зимостойкости хвои сосны благодаря синтезу в ней масел, способных повышать эластичность цитоплазмы и понижать температуру ее замерзания.
Обнаружены различия в ферментативных системах у водных и полупогруженных, болотных и наземных растений. В листьях водных растений преобладают гликолитические ферменты, а в листьях наземных — ферменты аэробного цикла. Особенно поразительно биохимическое и экологическое сходство в дыхании болотных растений с ныряющими животными (Р. М. Кроуфорд, 1981).
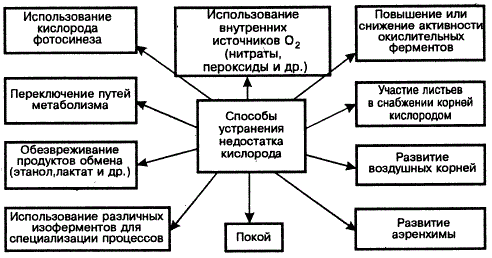
Высшие растения, как правило, обеспечивают свои потребности в кислороде не с помощью специальной системы, транспортирующей кислород, а путем непосредственного газообмена со средой, что возможно при достаточно развитой поверхности тела, обилии устьиц и межклеточных пространств. Несмотря на это, в природной обстановке нередко ограничивается дыхание как у суккулентов, покрытых толстой кутикулой, пробкой и восковым налетом, так и у водных растений, а также при затоплении и орошении, на заболоченных и плотных почвах. Разные виды растений неодинаково устойчивы к анаэробиозу (D. D. Hook, Р М. Crawford, 1978). Существуют приспособления к анаэробиозу, например система воздухоносных полостей (болотные и водные растения), по которым воздух проходит из листьев в корни и обратно. При этом листья выполняют защитную роль как на свету, так и в темноте; В этом есть необходимость. Так, содержание O2 в зарослях Menyathes trifoliata на глубине 25 см равно 0,47%, а на глубине 30—35 см в зарослях Equisetum limosum — 0,34%. Воздух к корням таких растений поступает только через надводные части (Б. Уильямс, Д. Барбер, 1964). Поэтому растения без аэренхимы обычно не могут выжить, если их корневые системы погружены в воду. Отсюда внутреннее строение водных растений напоминает сложные пчелиные соты с толщиной стенок в одну клетку, снабженных мелкими порами для обеспечения газом. Стебли водных растений обычно осуществляют резервную и транспортную функцию в доставке корням O2.
Как известно, у водных растений основная масса тела находится под водой в условиях плохой аэрации. В анаэробной среде выживание обеспечивается блокированием накопления токсических веществ в корнях (этанол, лактат и малат). Однако не только биохимические изменения обусловливают выживание организмов и анаэробной среде, важны и морфологические приспособления: увеличение объема межклеточных пространств, облегчающих диффузию газа при приливах и отливах воды между погруженными и непогруженными частями растения. На примере осоки (гигрофит) показано, что ее метаболизм в анаэробной среде неустойчив, при этом наблюдается даже деструкция мембран (Б. Б. Вартапетян, 1982). Отсюда сделан вывод, что устойчивость гигрофитов к отсутствию кислорода обусловлена не биохимическими механизмами (переключение обмена на альтернативный путь), а анатомо-физиологическими изменениями на уровне целого организма, т. е. развитием способности транспортировать O2 из надземной части к подземной. В этой связи автор предлагает отнести гигрофиты к растениям, обладающим не истинной, а кажущейся устойчивостью к отсутствию кислорода. Данные об использовании разных механизмов для адаптации растений к анаэробиозу свидетельствуют о взаимообусловленности изменений формы и функций в эволюции.
Лишение клетки O2 резко сказывается на ее энергетике, структуре и функции митохондрий. При кратковременной аноксии происходят разбухание органелл, сокращение числа крист, деградация митохондриальных мембран. В отсутствие экзогенной глюкозы из-за энергетического голодания аноксия приводит к деструкции ультраструктуры клеток. Интересна отметить, что клетки корней взрослых растений риса и осоки, живущих на переувлажненных анаэробных почвах, не обладают большей резистентностью к аноксии, чем корни тыквы и других растений (обитающих на хорошо аэрируемых почвах). Адаптация к анаэробиозу выступает как свойство целого растения. Из надземной части к корням поступает лишь незначительная часть O2. Адаптация к гипоксии и аноксии может быть обусловлена изменениями как на молекулярном, так и на уровне целого организма. У истинно устойчивых к аноксии растений есть молекулярные механизмы резистентности. У кажущихся устойчивыми растений нет таких механизмов, однако они способны жить в анаэробных условиях благодаря транспорту O2 из аэрируемых органов (Б. Б. Вартапетян, 1985).
Метаболические пути приспособления растений к гипо- и аноксии разнообразны, и на разных их уровнях осуществляется система регуляции (Т. В. Чиркова, 1988). С усилением напряженности анаэробиоза доминирующими становятся метаболическая регуляция, комплекс изменений для поддержания гомеостаза и репаративных процессов. Ими затрагиваются ЭТЦ митохондрий, осуществляются «сброс» восстановительных пиридиннуклеотидов, трансформация дыхательных путей, возрастание доли ПФП и синтез белков-ферментов анаэробного обмена, перестройка мембран, устраняется накопление токсических продуктов метаболизма (лактат, этанол и др.).
Опытами установлено, что при помещении корней растений в анаэробные условия листья их поглощают больше кислорода, чем в аэробной среде. У гигрофитов (манник) даже в нормальных условиях кислородного снабжения корней листья поглощают больше кислорода, чем требуется им самим. Как указывалось, роль листьев в снабжении растений кислородом очень велика, поэтому при лишении их листьев и света растения обнаруживают все симптомы анаэробного обмена: Накопление молочной кислоты, снижение дыхательного коэффициента.
В условиях, затрудняющих снабжение корней кислородом через листья (например, под водой или ледяной коркой), растения обогащаются органическими кислотами, свободными аминокислотами, при этом меняется ультраструктура клеток корня, повреждаются митохондрии, цистерны эндоплазматического ретикулума, уменьшаются количество рибосом в цитоплазме и размеры ядра. Такие изменения в клетках менее выражены у устойчивых к анаэробиозу видов (болотные растения).
В условиях анаэробиоза в результате комплекса внутриклеточных процессов происходит угнетение роста и развития, продуктивности растений. Как показывают исследования Г. М. Гриневой (1974), во избежание этого осуществляются перераспределение кислорода между органами растений, саморегуляция энергетического обмена путем переключения путей метаболизма в клетке, в том числе накопление ингибиторов роста и недоокисленных промежуточных продуктов (углеводы, спирты, органические кислоты), ненасыщенных липидов (выполняющих роли акцепторов водорода и заменителей кислорода), выделение токсичных веществ корнями и т. д. Адаптация растений к анаэробиозу связана с повышением активности цитохромоксидаз (что поддерживает межмембранный транспорт электронов); накоплением быстро утилизирующихся веществ (сукцинат и малат); усилением спиртового брожения (устраняющего токсическое накопление пирувата и ацетальдегида), нитратного дыхания (при этом устраняется токсическое накопление нитратов) и окислением глюкозы через ПФП; повышением роли гликолиза в синтезе АТФ (проявление эволюционно древнего способа энергетики в онтогенезе): Для предотвращения отрицательного влияния анаэробиоза имеет значение также образование дополнительных корней и газовых полостей у основания побега.
Приспособление растений выражается и в различной устойчивости к понижению или повышению температуры. Обычно оптимум температуры для дыхания у высокогорных и полярных растений лежит значительно ниже, чем у растений умеренной зоны, пустынь и тропиков. Они заметно различаются и по компенсационной точке фотосинтеза. У указанных растений меняется и качество ферментов, определяемое по константам его действия (А. В. Благовещенский, 1966). В целом у зимостойких видов интенсивность дыхания зимой выше, чем у незимостойких. У первых при понижении температуры наблюдается усиление окислительно-восстановительных процессов. У растений, произрастающих в крайних условиях существования, качество ферментов высокое, что дает им возможность сохранить активность химических реакций при резкой смене условий. Повышение качества ферментов идет параллельно увеличению доли низкомолекулярных белков — альбуминов и глобулинов. Такой параллелизм наблюдается как в онтогенезе, так и в филогенезе.
В целом адаптация растений к условиям обитания сопряжена с изменениями в метаболизме, энергетическими затратами. Как показывают многие наблюдения, в неблагоприятных условиях изменения в фотосинтезе оказываются более значительными, чем в дыхании. Это создает нехватку энергетического материала для роста и морфогенеза, включения регенерационных процессов и механизмов осмотических адаптаций, ведет к снижению репродуктивного потенциала растений. В итоге наблюдается общее истощение индивидуума и потеря устойчивости к стрессам. Поддержание стабильности растений в неблагоприятных условиях связано с усилением затрат на дыхание («дыхательная цена»). Сами эти затраты можно определить количеством израсходованного углерода, и они оказываются неравноценными у адаптированных и не адаптированных к стрессам растений (О. А. Семихатова, 1995). Уровень ингибирования растений определяется разницей между накоплением (фотосинтез) и расходованием (дыхание) энергетических ресурсов в неблагоприятных условиях. Все это делает чрезвычайно важным учет энергетических затрат при оценке адаптации растений и ведет к усилению давления отбора на их уменьшение при одновременном обеспечении репродукционного успеха индивидуума.
Одним из примеров экологических различий в дыхании растений является фотодыхание.