После того, как было доказано явление кроссинговера генетическими методами, необходимо было получить прямое доказательство обмена участками гомологичных хромосом, сопровождающегося рекомбинацией генов.
Это удалось сделать цитогенетическим методом в начале 30-х годов К. Штерну на дрозофиле и Б. Мак-Клинток и Г. Крейтон на кукурузе.
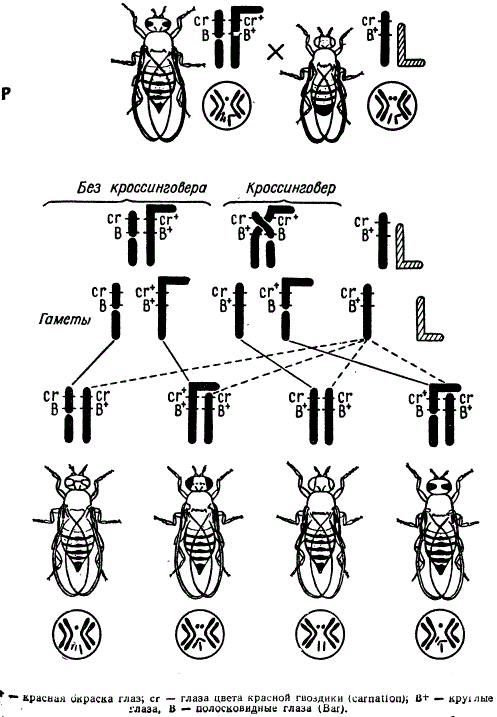
Цитологическое доказательство кроссинговера у дрозофилы
К. Штерну удалось получить у самки и у самца половые хромосомы, отличимые друг от друга цитологически и генетически, различающиеся по морфологии. У самок отличие Х-хромосом получено за счет того, что к одной из Х-хромосом был присоединен фрагмент от Y-хромосомы, что дало г-образную Х-хромосому с генами cr+ В+ дикого типа. Другая Х-хромосома была как бы составной: один из ее фрагментов, несущий центромеру, содержал рецессивный ген cr и доминантный ген В (ген cr определяет окраску глаза цвета красной гвоздики — carnation, а доминантный к дикому типу ген В — полосковидные глаза). Бесцентромерный фрагмент Х-хромосомы не был потерян в силу того, что он прикрепился к IV (маленькой) хромосоме. Так как прибавка фрагмента от Y-хромосомы, генетически мало активной, в общем не изменила хромосомного баланса зиготы, гетерозиготная по данным генам самка была вполне жизнеспособна. В гаметогенезе у такой самки на участке между указанными генами мог происходить кроссинговер, который учитывался в потомстве как цитологически, так и генетически.
Для анализирующего скрещивания были взяты самцы с нормальными X — и Y-хромосомами, при этом половая хромосома самца несла гены cr и В+ в гемизиготном состоянии. В потомстве были изучены цитологически только самки, среди которых возникли два класса мух с некроссоверными хромосомами: cr B/cr B+ и cr+B+/cr B+, и два других класса мух: cr B+/cr B+ и cr+B/cr B+.
Эти два последних класса самок могли появиться только при условии произошедшего кроссинговера между генами cr и B. И так как каждый из четырех классов отличался друг от друга не только по признакам глаз, но и по морфологии половых хромосом, то цитологически можно было установить, что кроссинговер, давший два новых класса мух, сопровождался обменом участками Х-хромосом.
К. Штерн проверил цитологически 374 самки из описанного скрещивания, 369 из них соответствовали ожидаемому, и лишь 5 самок были исключением, причина появления которого не была выяснена.
Подобный же опыт был проведен на кукурузе. Г. Крейтон и Б. Мак-Клинток удалось пометить хромосомы одной пары генетически и морфологически и получить линию кукурузы, у которой IX пара хромосом оказалась гетероморфной — одна была нормальной, а другая несла утолщение на конце одного плеча, в то время как другое ее плечо было заметно удлинено по сравнению с таковым у нормальной IX хромосомы. Эти особенности в строении гетероморфной пары хромосом прекрасно обнаруживались при цитологических исследованиях.
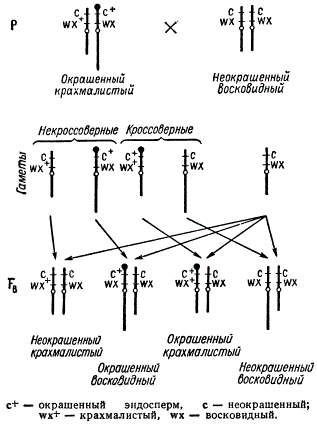
Цитологическое доказательство кроссинговера у кукурузы
В опыте нормальная хромосома несла рецессивный ген с (неокрашенный эндосперм) и доминантный ген wx+ (крахмалистый эндосперм). Измененная хромосома несла доминантный ген с+ (окрашенный эндосперм) и рецессивный ген wx (восковидный эндосперм). Дигетерозиготу с+ wx/c wx+ скрещивали с линией, имеющей морфологически нормальные хромосомы с рецессивными генами с и wx: c+ wx/c wx+ x c wx/c wx. В потомстве получили как некроссоверные, так и кроссоверные зерна. При цитологическом изучении их было обнаружено, что кроссоверные зерна неизменно содержали IX хромосому с обменявшимися участками: хромосому нормальной длины, но с утолщением или хромосому без утолщения, но удлиненную.
Таким образом, одновременно цитологически и генетически было показано, что рекомбинация генов сопровождается обменом участками гомологичных хромосом. После этого кроссинговер перестал быть гипотезой, а стал реально доказанным явлением, но механизм его остается до сих пор не выясненным окончательно.
Итак, изучение сцепления и перекреста хромосом генетическими методами позволило прийти к следующим важным выводам.
1. В хромосоме может находиться более чем один ген.
2. Гены, находящиеся в одной хромосоме, в случае полного сцепления наследуются в потомстве гибрида как одна аллельная пара.
3. Обычно сцепление нарушается кроссинговером; кроссинговер является регулярным, нормально протекающим в мейозе процессом обмена идентичными участками гомологичных хромосом.
4. Кроссинговер приводит к рекомбинации сцепленных генов, благодаря чему значительно расширяется возможность комбинативной изменчивости, служащей важным источником для естественного отбора и селекции.
5. Величина кроссинговера (процент рекомбинаций) зависит от расстояния между рекомбинирующимися генами. Процент рекомбинаций может колебаться от долей процента до 50%, но никогда не достигает этого значения.
6. Кроссинговер может быть одинарным, двойным и множественным.
7. Частота появления рекомбинантных потомков от одинарного обмена всегда превышает таковую от двойных и множественных обменов.
8. Для двух крайних генов А и С из трех (АВС) частота кроссинговера меньше суммы одинарных кроссинговеров между генами А и В и между В и С.
9. Вследствие явления интерференции один разрыв (обмен) препятствует осуществлению других обменов на ближайшем участке хромосомы. С увеличением расстояния интерференция уменьшается.
В результате изучения кроссинговера была доказана наследственная дискретность хромосом, сформулирована теория гена и Установлено линейное расположение генов в хромосоме. Исходя из этой теории, были установлены группы сцепления генов и построены генетические карты хромосом, количество которых для изученных видов совпадает с гаплоидным числом хромосом.
Явление кроссинговера было вначале показано на основе генетического анализа, а затем получило полное подтверждение в цитологических исследованиях.